
Characterization of Marine Organism Extracellular Matrix-Anchored Extracellular Vesicles and Their Biological Effect on the Alleviation of Pro-Inflammatory Cytokines


Abstract
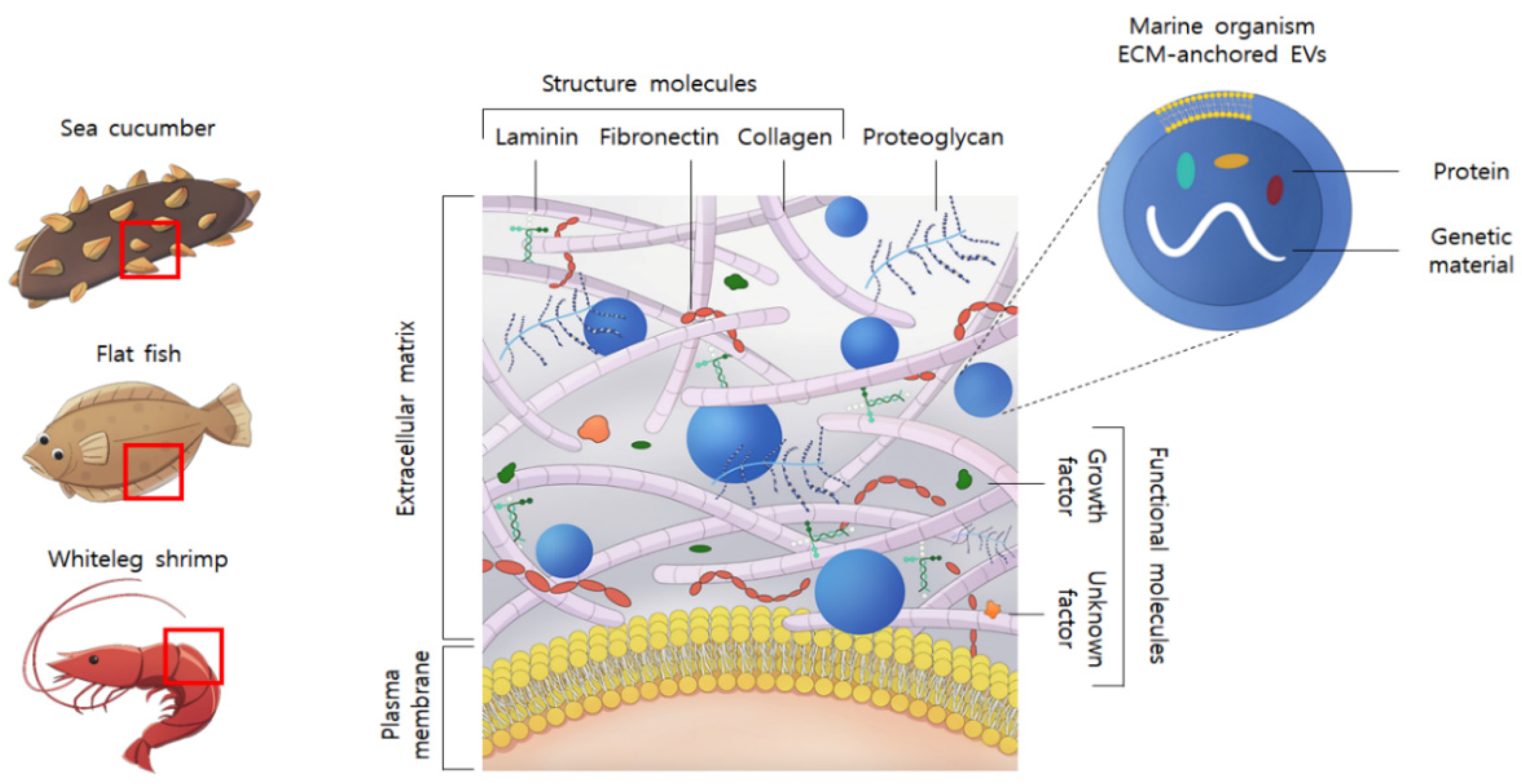
:1. Introduction
2. Results and Discussion
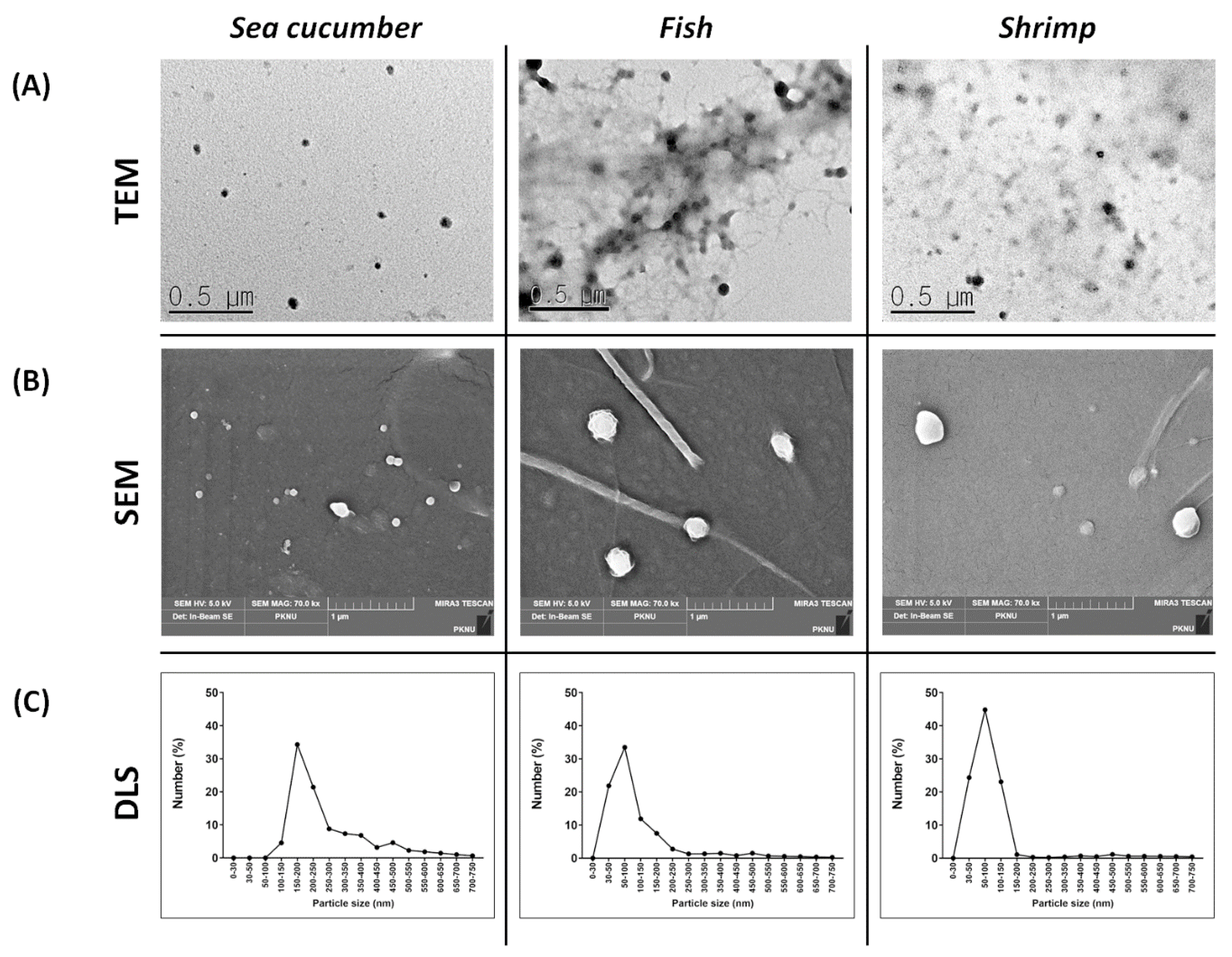
2.1. Physical Characterization of the Marine Organism-Derived EVs
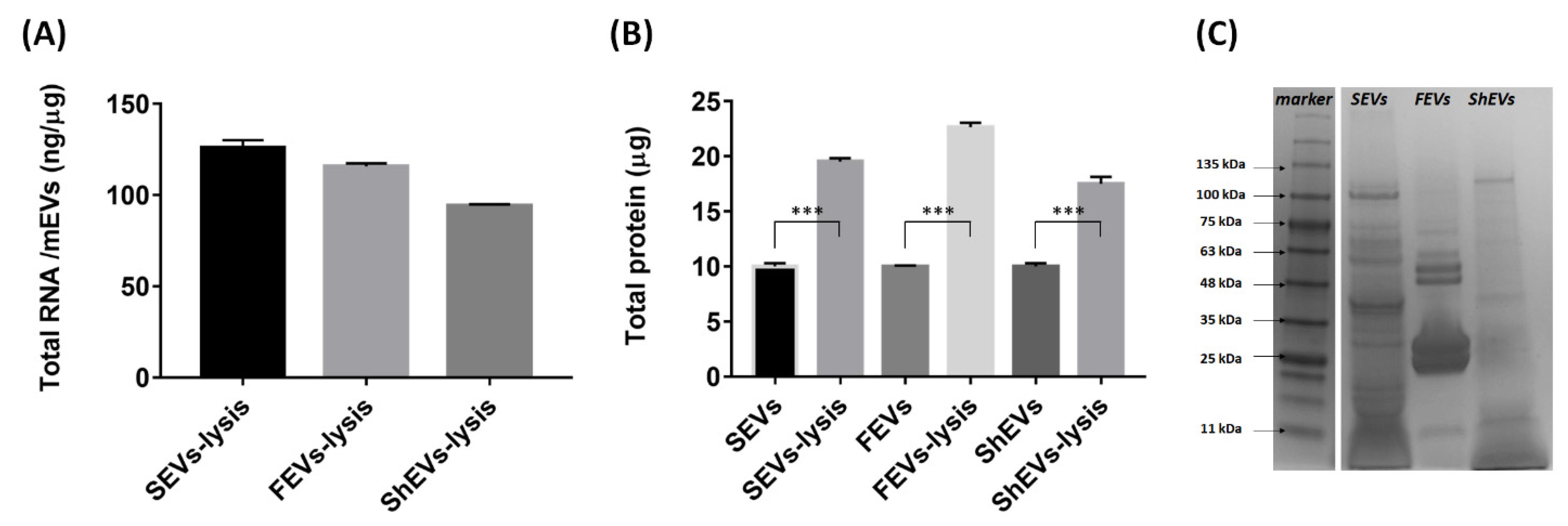
2.2. Marine Organism-Derived EVs Contain Genetic Material and Protein
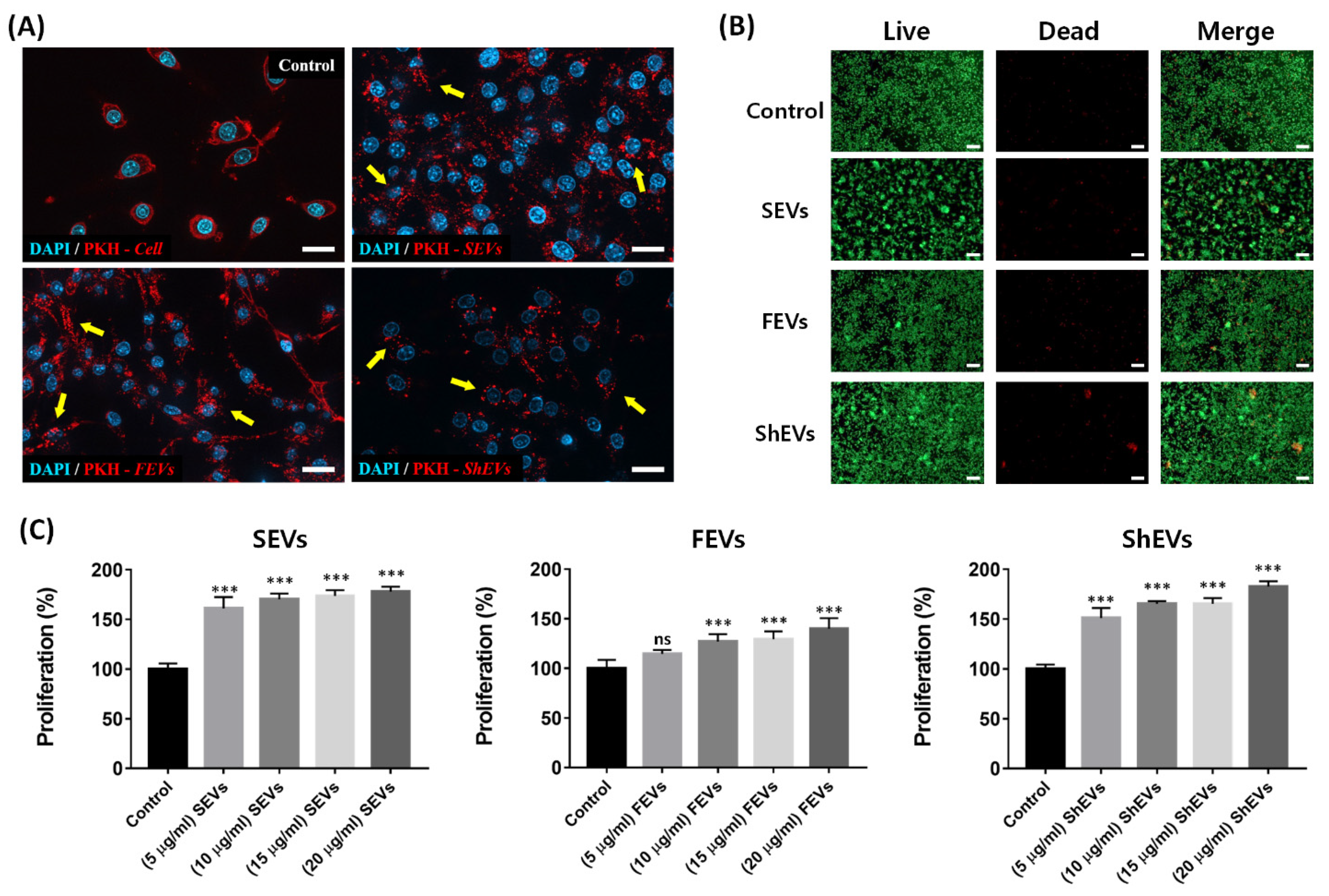
2.3. Cytotoxicity Analysis
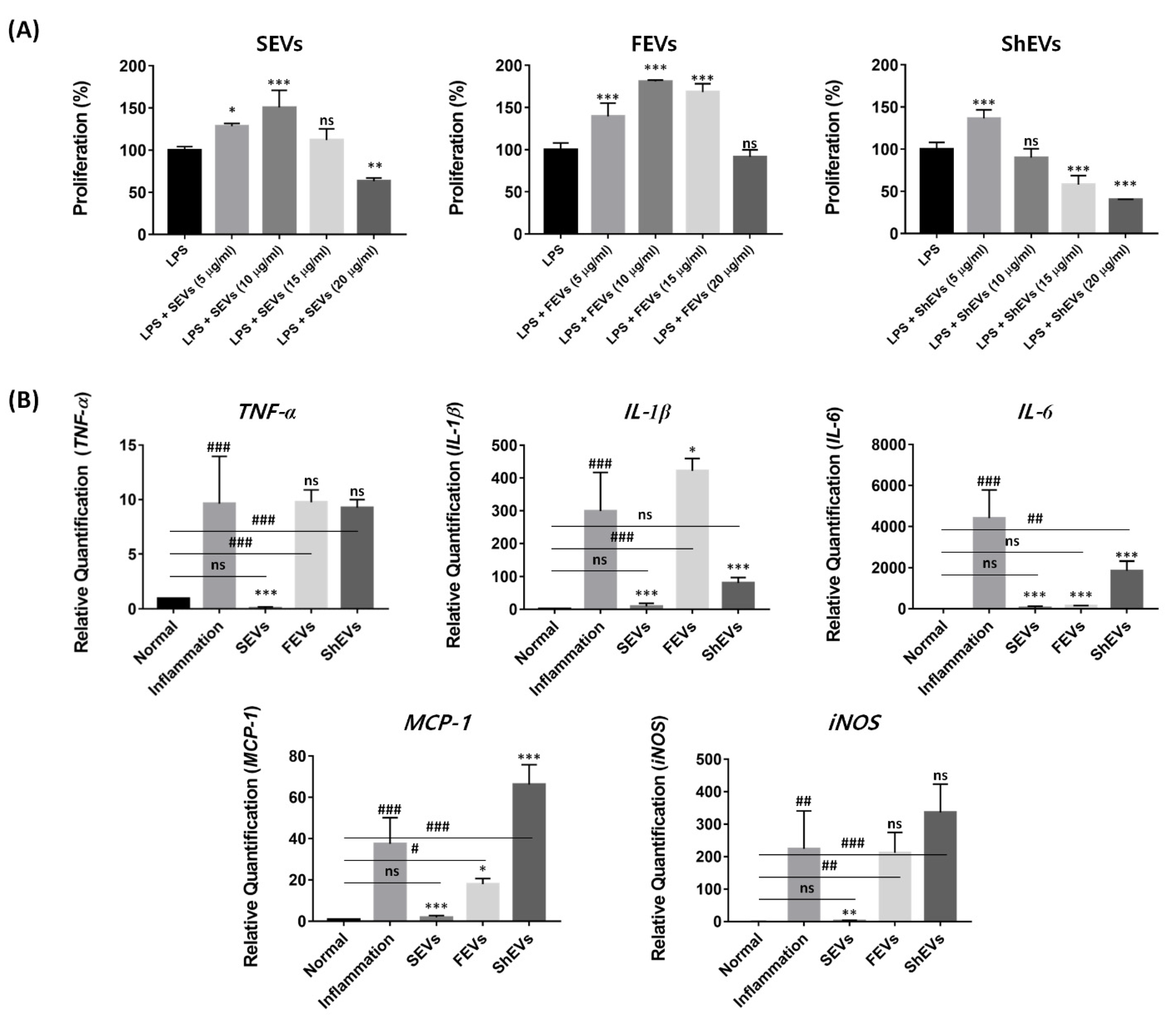
2.4. Modulation of Pro-Inflammatory Cytokine
3. Materials and Methods
3.1. Preparation of Marine EVs
3.2. Marine EV Morphology
3.3. Particle Size Analysis
3.4. Cell Culture
3.5. Immune Modulation Assay
3.6. Fluorescence Microscopy
3.7. Protein Cargo Analysis
3.8. Gene Cargo and Gene Expression Analysis
3.9. Assessment of Cell Viability and Proliferation
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kim, S.-K. Marine Biomaterials: Characterization, Isolation and Applications; CRC Press: Boca Raton, FL, USA, 2013; ISBN 1466505648. [Google Scholar]
- Rahman, M. Collagen of Extracellular Matrix from Marine Invertebrates and Its Medical Applications. Mar. Drugs 2019, 17, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seixas, M.J.; Martins, E.; Reis, R.L.; Silva, T.H. Extraction and Characterization of Collagen from Elasmobranch Byproducts for Potential Biomaterial Use. Mar. Drugs 2020, 18, 617. [Google Scholar] [CrossRef]
- Zhong, M.; Chen, T.; Hu, C.; Ren, C. Isolation and Characterization of Collagen from the Body Wall of Sea Cucumber Stichopus Monotuberculatus. J. Food Sci. 2015, 80, C671–C679. [Google Scholar] [CrossRef] [PubMed]
- Badylak, S.F. Xenogeneic Extracellular Matrix as a Scaffold for Tissue Reconstruction. Transpl. Immunol. 2004, 12, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Hodde, J.P.; Record, R.D.; Liang, H.A.; Badylak, S.F. Vascular Endothelial Growth Factor in Porcine-Derived Extracellular Matrix. Endothelium 2001, 8, 11–24. [Google Scholar] [CrossRef]
- Agrawal, V.; Tottey, S.; Johnson, S.A.; Freund, J.M.; Siu, B.F.; Badylak, S.F. Recruitment of Progenitor Cells by an Extracellular Matrix Cryptic Peptide in a Mouse Model of Digit Amputation. Tissue Eng. Part A 2011, 17, 2435–2443. [Google Scholar] [CrossRef] [Green Version]
- Sicari, B.M.; Dziki, J.L.; Siu, B.F.; Medberry, C.J.; Dearth, C.L.; Badylak, S.F. The Promotion of a Constructive Macrophage Phenotype by Solubilized Extracellular Matrix. Biomaterials 2014, 35, 8605–8612. [Google Scholar] [CrossRef]
- Huleihel, L.; Hussey, G.S.; Naranjo, J.D.; Zhang, L.; Dziki, J.L.; Turner, N.J.; Stolz, D.B.; Badylak, S.F. Matrix-Bound Nanovesicles within ECM Bioscaffolds. Sci. Adv. 2016, 2, e1600502. [Google Scholar] [CrossRef] [Green Version]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Bedina Zavec, A.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological Properties of Extracellular Vesicles and Their Physiological Functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [Green Version]
- Meldolesi, J. Exosomes and Ectosomes in Intercellular Communication. Curr. Biol. 2018, 28, R435–R444. [Google Scholar] [CrossRef] [Green Version]
- Rekker, K.; Saare, M.; Roost, A.M.; Kubo, A.-L.; Zarovni, N.; Chiesi, A.; Salumets, A.; Peters, M. Comparison of Serum Exosome Isolation Methods for MicroRNA Profiling. Clin. Biochem. 2014, 47, 135–138. [Google Scholar] [CrossRef]
- Dutta, D.; Khan, N.; Wu, J.; Jay, S.M. Extracellular Vesicles as an Emerging Frontier in Spinal Cord Injury Pathobiology and Therapy. Trends Neurosci. 2021, 44, 492–506. [Google Scholar] [CrossRef]
- Barreiro, K.; Dwivedi, O.P.; Leparc, G.; Rolser, M.; Delic, D.; Forsblom, C.; Groop, P.; Groop, L.; Huber, T.B.; Puhka, M.; et al. Comparison of Urinary Extracellular Vesicle Isolation Methods for Transcriptomic Biomarker Research in Diabetic Kidney Disease. J. Extracell. Vesicles 2020, 10, e12038. [Google Scholar] [CrossRef] [PubMed]
- Perge, P.; Butz, H.; Pezzani, R.; Bancos, I.; Nagy, Z.; Pálóczi, K.; Nyírő, G.; Decmann, Á.; Pap, E.; Luconi, M.; et al. Evaluation and Diagnostic Potential of Circulating Extracellular Vesicle-Associated MicroRNAs in Adrenocortical Tumors. Sci. Rep. 2017, 7, 5474. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, A.; Kim, H.S.; Bojmar, L.; Gyan, K.E.; Cioffi, M.; Hernandez, J.; Zambirinis, C.P.; Rodrigues, G.; Molina, H.; Heissel, S.; et al. Extracellular Vesicle and Particle Biomarkers Define Multiple Human Cancers. Cell 2020, 182, 1044–1061. [Google Scholar] [CrossRef]
- de Cassia Noronha, N.; Mizukami, A.; Caliári-Oliveira, C.; Cominal, J.G.; Rocha, J.L.M.; Covas, D.T.; Swiech, K.; Malmegrim, K.C.R. Priming Approaches to Improve the Efficacy of Mesenchymal Stromal Cell-Based Therapies. Stem Cell Res. Ther. 2019, 10, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Chopp, M. Exosome Therapy for Stroke. Stroke 2018, 49, 1083–1090. [Google Scholar] [CrossRef]
- Ni, Z.; Zhou, S.; Li, S.; Kuang, L.; Chen, H.; Luo, X.; Ouyang, J.; He, M.; Du, X.; Chen, L. Exosomes: Roles and Therapeutic Potential in Osteoarthritis. Bone Res. 2020, 8, 1–18. [Google Scholar] [CrossRef]
- Gilligan, K.E.; Dwyer, R.M. Engineering Exosomes for Cancer Therapy. Int. J. Mol. Sci. 2017, 18, 1122. [Google Scholar] [CrossRef] [Green Version]
- Andaloussi, S.E.; Mäger, I.; Breakefield, X.O.; Wood, M.J. Extracellular Vesicles: Biology and Emerging Therapeutic Opportunities. Nat. Rev. Drug Discov. 2013, 12, 347–357. [Google Scholar] [CrossRef]
- Huleihel, L.; Bartolacci, J.G.; Dziki, J.L.; Vorobyov, T.; Arnold, B.; Scarritt, M.E.; Pineda Molina, C.; LoPresti, S.T.; Brown, B.N.; Naranjo, J.D. Matrix-Bound Nanovesicles Recapitulate Extracellular Matrix Effects on Macrophage Phenotype. Tissue Eng. Part A 2017, 23, 1283–1294. [Google Scholar] [CrossRef] [PubMed]
- An, M.; Kwon, K.; Park, J.; Ryu, D.-R.; Shin, J.-A.; Kang, J.L.; Choi, J.H.; Park, E.-M.; Lee, K.E.; Woo, M. Extracellular Matrix-Derived Extracellular Vesicles Promote Cardiomyocyte Growth and Electrical Activity in Engineered Cardiac Atria. Biomaterials 2017, 146, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Biller, S.J.; Schubotz, F.; Roggensack, S.E.; Thompson, A.W.; Summons, R.E.; Chisholm, S.W. Bacterial Vesicles in Marine Ecosystems. Science 2014, 343, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Scanlan, D. Bacterial Vesicles in the Ocean. Science 2014, 343, 143–144. [Google Scholar] [CrossRef]
- Jo, S.-H.; Kim, C.; Park, S.-H. Novel Marine Organism-Derived Extracellular Vesicles for Control of Anti-Inflammation. Tissue Eng. Regen. Med. 2021, 18, 71–79. [Google Scholar] [CrossRef]
- Sunkara, V.; Woo, H.-K.; Cho, Y.-K. Emerging Techniques in the Isolation and Characterization of Extracellular Vesicles and Their Roles in Cancer Diagnostics and Prognostics. Analyst 2016, 141, 371–381. [Google Scholar] [CrossRef] [Green Version]
- Szatanek, R.; Baj-Krzyworzeka, M.; Zimoch, J.; Lekka, M.; Siedlar, M.; Baran, J. The Methods of Choice for Extracellular Vesicles (EVs) Characterization. Int. J. Mol. Sci. 2017, 18, 1153. [Google Scholar] [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular Vesicles: Exosomes, Microvesicles, and Friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Shah, R.; Patel, T.; Freedman, J.E. Circulating Extracellular Vesicles in Human Disease. N. Engl. J. Med. 2018, 379, 958–966. [Google Scholar] [CrossRef]
- Kim, M.; Shin, D.I.; Choi, B.H.; Min, B.-H. Exosomes from IL-1β-Primed Mesenchymal Stem Cells Inhibited IL-1β- and TNF-α-Mediated Inflammatory Responses in Osteoarthritic SW982 Cells. Tissue Eng. Regen. Med. 2021, 18, 525–536. [Google Scholar] [CrossRef]
- Weng, Y.; Sui, Z.; Shan, Y.; Hu, Y.; Chen, Y.; Zhang, L.; Zhang, Y. Effective Isolation of Exosomes with Polyethylene Glycol from Cell Culture Supernatant for In-Depth Proteome Profiling. Analyst 2016, 141, 4640–4646. [Google Scholar] [CrossRef]
- Park, S.-Y.; Lim, H.K.; Lee, S.; Hwang, H.C.; Cho, S.K.; Cho, M. Pepsin-Solubilised Collagen (PSC) from Red Sea Cucumber (Stichopus Japonicus) Regulates Cell Cycle and the Fibronectin Synthesis in HaCaT Cell Migration. Food Chem. 2012, 132, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Benjakul, S.; Nalinanon, S.; Shahidi, F. Fish Collagen. In Food Biochemistry and Food Processing; Simpson, B.K., Ed.; Wiley-Blackwell: Oxford, UK, 2012; pp. 365–387. ISBN 9781118308035. [Google Scholar]
- Sriket, P.; Benjakul, S.; Visessanguan, W.; Kijroongrojana, K. Comparative Studies on Chemical Composition and Thermal Properties of Black Tiger Shrimp (Penaeus Monodon) and White Shrimp (Penaeus Vannamei) Meats. Food Chem. 2007, 103, 1199–1207. [Google Scholar] [CrossRef]
- Van der Pol, E.; Böing, A.N.; Harrison, P.; Sturk, A.; Nieuwland, R. Classification, Functions, and Clinical Relevance of Extracellular Vesicles. Pharmacol. Rev. 2012, 64, 676–705. [Google Scholar] [PubMed] [Green Version]
- Chuo, S.T.-Y.; Chien, J.C.-Y.; Lai, C.P.-K. Imaging Extracellular Vesicles: Current and Emerging Methods. J. Biomed. Sci. 2018, 25, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Feng, D.; Zhao, W.-L.; Ye, Y.-Y.; Bai, X.-C.; Liu, R.-Q.; Chang, L.-F.; Zhou, Q.; Sui, S.-F. Cellular Internalization of Exosomes Occurs through Phagocytosis. Traffic 2010, 11, 675–687. [Google Scholar] [CrossRef]
- Orekhov, A.N.; Orekhova, V.A.; Nikiforov, N.G.; Myasoedova, V.A.; Grechko, A.V.; Romanenko, E.B.; Zhang, D.; Chistiakov, D.A. Monocyte Differentiation and Macrophage Polarization. Vessel Plus 2019, 3, 10. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The Chemokine System in Diverse Forms of Macrophage Activation and Polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Park, D.Y.; Yun, H.-W.; Lim, S.; Truong, M.-D.; Yin, X.Y.; Park, J.; Kim, B.K.; Shin, D.I.; Li, X.G.; Chung, J.Y. Cross-linked Cartilage Acellular Matrix Film Decreases Postsurgical Peritendinous Adhesions. Artif. Organs 2020, 44, E136–E149. [Google Scholar] [CrossRef]
- Hellström, M.; El-Akouri, R.R.; Sihlbom, C.; Olsson, B.M.; Lengqvist, J.; Bäckdahl, H.; Johansson, B.R.; Olausson, M.; Sumitran-Holgersson, S.; Brännström, M. Towards the Development of a Bioengineered Uterus: Comparison of Different Protocols for Rat Uterus Decellularization. Acta Biomater. 2014, 10, 5034–5042. [Google Scholar] [CrossRef]
- Yoshimasa, Y.; Maruyama, T. Bioengineering of the Uterus. Reprod. Sci. 2021, 28, 1596–1611. [Google Scholar] [CrossRef] [PubMed]
- Gattazzo, F.; Urciuolo, A.; Bonaldo, P. Extracellular Matrix: A Dynamic Microenvironment for Stem Cell Niche. Biochim. Biophys. Acta 2014, 1840, 2506–2519. [Google Scholar] [CrossRef] [PubMed]
- Siahaan, E.A.; Pangestuti, R.; Munandar, H.; Kim, S.-K. Cosmeceuticals Properties of Sea Cucumbers: Prospects and Trends. Cosmetics 2017, 4, 26. [Google Scholar] [CrossRef] [Green Version]
- Himaya, S.W.A.; Ryu, B.; Qian, Z.-J.; Kim, S.-K. Sea Cucumber, Stichopus Japonicus Ethyl Acetate Fraction Modulates the Lipopolysaccharide Induced INOS and COX-2 via MAPK Signaling Pathway in Murine Macrophages. Environ. Toxicol. Pharmacol. 2010, 30, 68–75. [Google Scholar] [CrossRef]
- Janakiram, N.B.; Mohammed, A.; Rao, C.V. Sea Cucumbers Metabolites as Potent Anti-Cancer Agents. Mar. Drugs 2015, 13, 2909–2923. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, M.; Lopes, I.; Teixeira, J.; Botelho, C.; Gomes, A.C. Exosome-like Nanoparticles: A New Type of Nanocarrier. Curr. Med. Chem. 2020, 27, 3888–3905. [Google Scholar] [CrossRef]
- Corradetti, B.; Gonzalez, D.; Pinto, I.M.; Conlan, R.S. Exosomes as Therapeutic Systems. Front. Cell Dev. Biol. 2021, 9, 714743. [Google Scholar] [CrossRef]
- Augst, A.D.; Kong, H.J.; Mooney, D.J. Alginate Hydrogels as Biomaterials. Macromol. Biosci. 2006, 6, 623–633. [Google Scholar] [CrossRef]
- Bellucci, D.; Sola, A.; Gazzarri, M.; Chiellini, F.; Cannillo, V. A New Hydroxyapatite-Based Biocomposite for Bone Replacement. Mater. Sci. Eng. C 2013, 33, 1091–1101. [Google Scholar] [CrossRef]
- Popowski, K.; Lutz, H.; Hu, S.; George, A.; Dinh, P.-U.; Cheng, K. Exosome Therapeutics for Lung Regenerative Medicine. J. Extracell. Vesicles 2020, 9, 1785161. [Google Scholar] [CrossRef]
- Ren, K. Exosomes in Perspective: A Potential Surrogate for Stem Cell Therapy. Odontology 2019, 107, 271–284. [Google Scholar] [CrossRef]
- Cai, Y.; Liu, W.; Lian, L.; Xu, Y.; Bai, X.; Xu, S.; Zhang, J. Stroke Treatment: Is Exosome Therapy Superior to Stem Cell Therapy? Biochimie 2020, 179, 190–204. [Google Scholar] [CrossRef] [PubMed]
- Carnevali, M.C. Regeneration in Echinoderms: Repair, Regrowth, Cloning. Invertebr. Surviv. J. 2006, 3, 64–76. [Google Scholar]
- Oh, G.-W.; Ko, S.-C.; Lee, D.H.; Heo, S.-J.; Jung, W.-K. Biological Activities and Biomedical Potential of Sea Cucumber (Stichopus Japonicus): A Review. Fish. Aquat. Sci. 2017, 20, 1–17. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Forward Sequences (5′–3′) | Reverse Sequences (5′–3′) |
|---|---|---|
| TNF-α | TTCTCATTCCTGCTTGTGGC | GGGAACTTCTCATCCCTTTGG |
| IL-1β | AAAGCTCTCCACCTCAATGG | GCCGTCTTTCATTACACAGG |
| IL-6 | GATACCACTCCCAACAGACC | GCAAGTGCATCATCGTTGTTC |
| MCP-1 | CCTGCTGCTACTCATTCACC | CTGGACCCATTCCTTCTTGG |
| iNOS | GTGGTGACAAGCACATTTGG | GAACTGAGGGTACATGCTGG |
| GAPDH | TTCAACAGCAACTCCCACTC | TCCTTGGAGGCCATGTAGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jo, S.-H.; Kim, S.-H.; Kim, C.; Park, S.-H. Characterization of Marine Organism Extracellular Matrix-Anchored Extracellular Vesicles and Their Biological Effect on the Alleviation of Pro-Inflammatory Cytokines. Mar. Drugs 2021, 19, 592. https://doi.org/10.3390/md19110592
Jo S-H, Kim S-H, Kim C, Park S-H. Characterization of Marine Organism Extracellular Matrix-Anchored Extracellular Vesicles and Their Biological Effect on the Alleviation of Pro-Inflammatory Cytokines. Marine Drugs. 2021; 19(11):592. https://doi.org/10.3390/md19110592
Chicago/Turabian StyleJo, Sung-Han, Seon-Hwa Kim, Changsu Kim, and Sang-Hyug Park. 2021. "Characterization of Marine Organism Extracellular Matrix-Anchored Extracellular Vesicles and Their Biological Effect on the Alleviation of Pro-Inflammatory Cytokines" Marine Drugs 19, no. 11: 592. https://doi.org/10.3390/md19110592