
Titanium Dioxide Nanoparticles Exacerbate Allergic Airway Inflammation via TXNIP Upregulation in a Mouse Model of Asthma

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Physicochemical Characterization of TiO2NPs
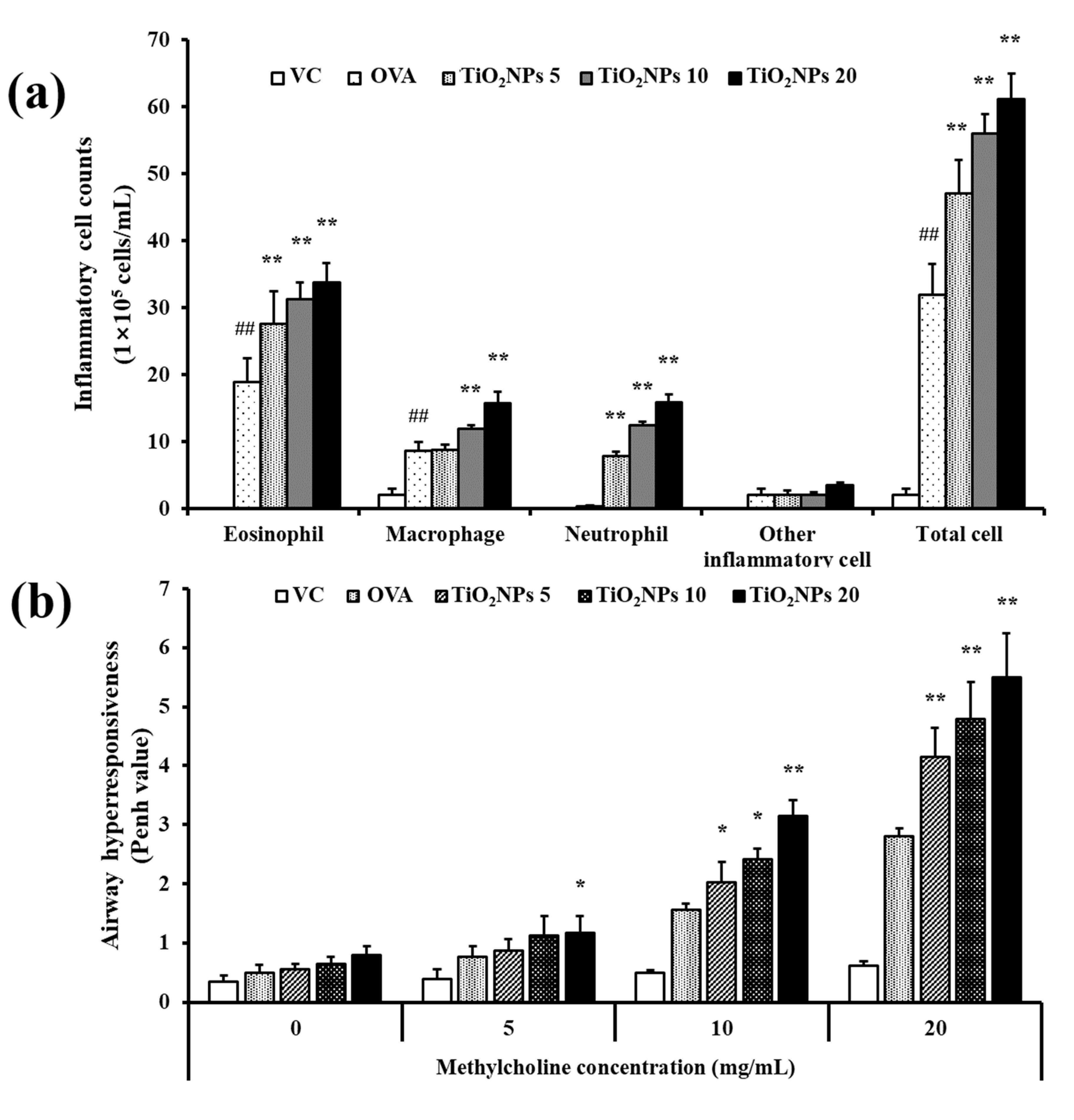
2.2. Effects of TiO2NPs on AHR and Inflammatory Cell Counts
2.3. Effects of TiO2NPs on Cytokine Production and OVA-Specific IgE Levels in Serum
2.4. Effects of TiO2NPs on Airway Inflammation and Mucous Secretion in OVA-Induced Mice
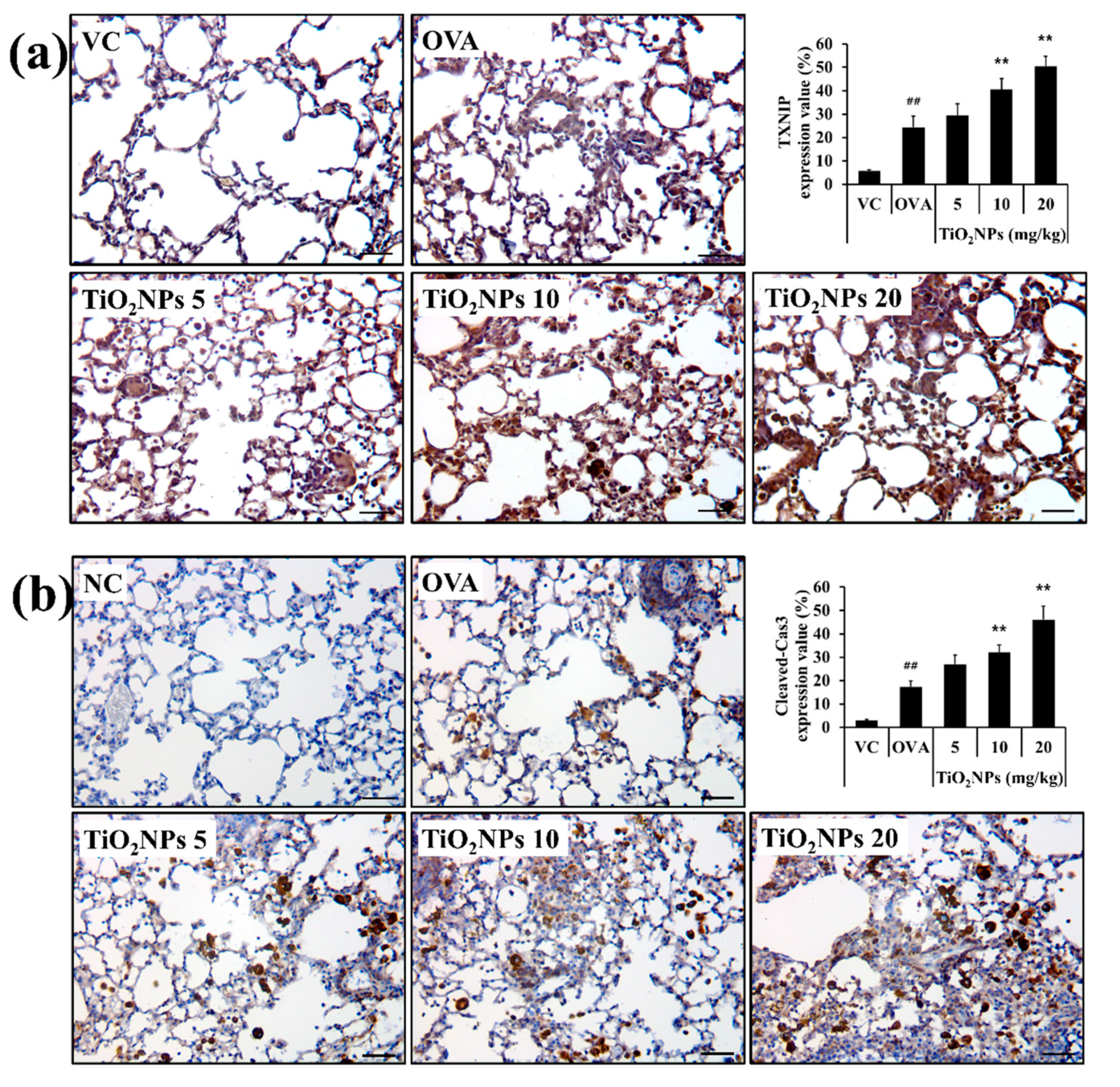
2.5. Effects of TiO2NPs on TXNIP and Apoptotic Protein Expression
2.6. Effects of TiO2NPs on the Production of Proinflammatory Mediators in NCI-H292 Cells
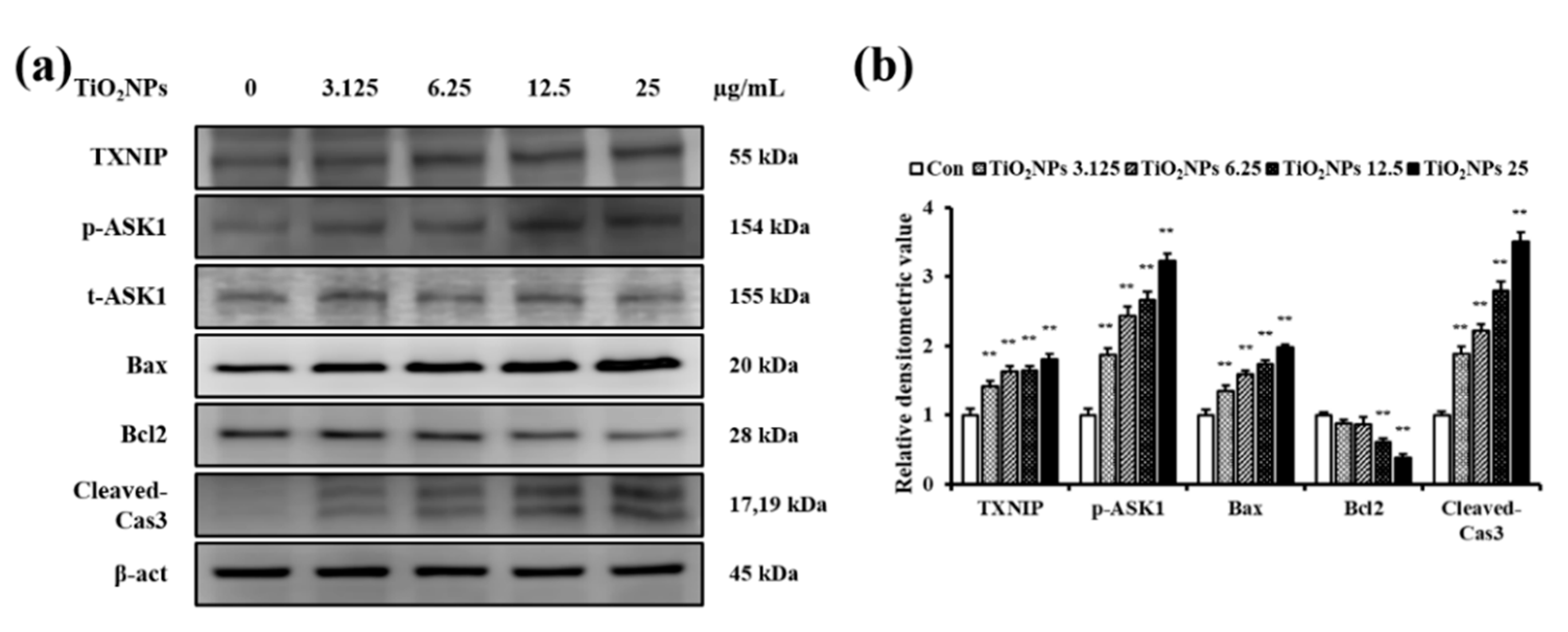
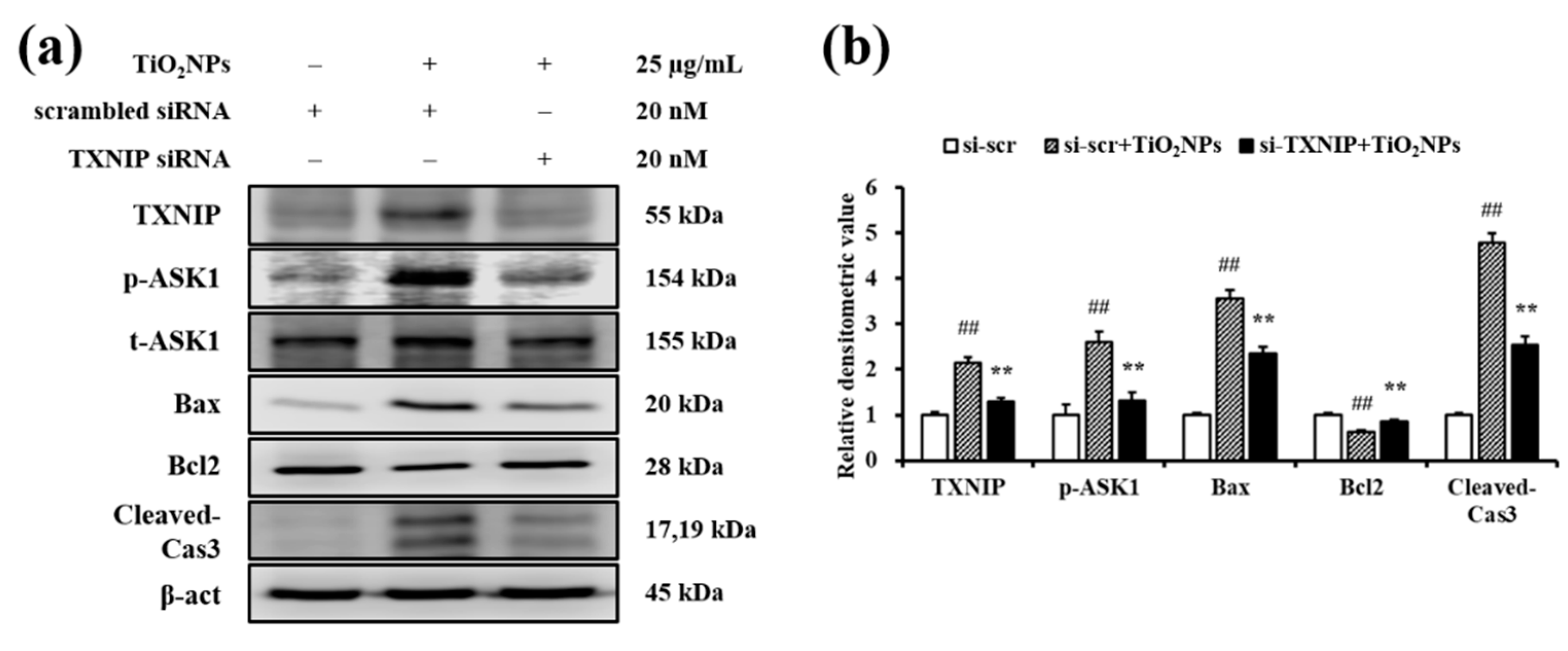
2.7. Effects of TiO2NPs on TXNIP and Apoptosis Protein Expression in NCI-H292 Cells
3. Discussion
4. Materials and Methods
4.1. Characterization of TiO2NPs
4.2. Experimental Procedure for Allergic Asthma Induction
4.3. Measurement of Airway AHR
4.4. Collection of Bronchoalveolar Lavage Fluid (BALF) and Cell Counting
4.5. Cytokines Assay
4.6. Histopathology and IHC
4.7. Western Blot Analysis
4.8. Cell Culture
4.9. Cell Viability Assay
4.10. Measurement of mRNA Expression of Proinflammatory Cytokines in NCI–H292 Cells
4.11. Small Interfering RNA Transfection of NCI-H292 Cells
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jonasson, S.; Gustafsson, A.; Koch, B.; Bucht, A. Inhalation exposure of nano-scaled titanium dioxide (TiO2) particles alters the inflammatory responses in asthmatic mice. Inhal. Toxicol. 2013, 25, 179–191. [Google Scholar] [CrossRef]
- Da Silva, A.L.; Cruz, F.F.; Rocco, P.R.M.; Morales, M.M. New perspectives in nanotherapeutics for chronic respiratory diseases. Biophys. Rev. 2017, 9, 793–803. [Google Scholar] [CrossRef] [Green Version]
- Ihrie, M.D.; Bonner, J.C. The toxicology of engineered nanomaterials in asthma. Curr. Environ. Health Rep. 2018, 5, 100–109. [Google Scholar] [CrossRef]
- Dasari, K.B.; Cho, H.; Jaćimović, R.; Sun, G.M.; Yim, Y.H. Chemical composition of asian dust in daejeon, korea, during the spring season. ACS Earth Space Chem. 2020, 4, 1227–1236. [Google Scholar] [CrossRef]
- Ambalavanan, N.; Stanishevsky, A.; Bulger, A.; Halloran, B.; Steele, C.; Vohra, Y.; Matalon, S. Titanium oxide nanoparticle instillation induces inflammation and inhibits lung development in mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2013, 304, L152–L161. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.G.; Lee, P.H.; Lee, S.H.; Park, M.K.; Jang, A.S. Effect of TiO2 nanoparticles on inflammasome-mediated airway inflammation and responsiveness. Allergy Asthma Immunol. Res. 2017, 9, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Frey, S.M.; Jones, M.R.; Goldstein, N.; Riekert, K.; Fagnano, M.; Halterman, J.S. Knowledge of inhaled therapy and responsibility for asthma management among young teens with uncontrolled persistent asthma. Acad. Pediatr. 2018, 18, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Alharris, E.; Alghetaa, H.; Seth, R.; Chatterjee, S.; Singh, N.P.; Nagarkatti, M.; Nagarkatti, P. Resveratrol attenuates allergic asthma and associated inflammation in the lungs through regulation of miRNA-34a that targets FoxP3 in mice. Front. Immunol. 2018, 9, 2992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menzel, M.; Ramu, S.; Calvén, J.; Olejnicka, B.; Sverrild, A.; Porsbjerg, C.; Tufvesson, E.; Bjermer, L.; Akbarshahi, H.; Uller, L. Oxidative stress attenuates TLR3 responsiveness and impairs anti-viral mechanisms in bronchial epithelial cells from COPD and asthma patients. Front. Immunol. 2019, 10, 2765. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Sim, S.; Park, H.S. Distinct functions of eosinophils in severe asthma with type 2 phenotype: Clinical implications. Korean J. Intern. Med. 2020, 35, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; DeKruyff, R.H.; Umetsu, D.T. The many paths to asthma: Phenotype shaped by innate and adaptive immunity. Nat. Immunol. 2010, 11, 577–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerner, A.G.; Upton, J.P.; Praveen, P.V.; Ghosh, R.; Nakagawa, Y.; Igbaria, A.; Shen, S.; Nguyen, V.; Backes, B.J.; Heiman, M.; et al. IRE1alpha induces thioredoxin-interacting protein to activate the NLRP3 inflammasome and promote programmed cell death under irremediable ER stress. Cell Metab. 2012, 16, 250–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelsaid, M.A.; Matragoon, S.; Ergul, A.; El-Remessy, A.B. Deletion of thioredoxin interacting protein (TXNIP) augments hyperoxia-induced vaso-obliteration in a mouse model of oxygen induced-retinopathy. PLoS ONE 2014, 9, e110388. [Google Scholar] [CrossRef]
- Mohamed, I.N.; Sarhan, N.R.; Eladl, M.A.; El-Remessy, A.B.; El-Sherbiny, M. Deletion of thioredoxin-interacting protein ameliorates high fat diet-induced non-alcoholic steatohepatitis through modulation of toll-like receptor 2-NLRP3-inflammasome axis: Histological and immunohistochemical study. Acta. Histochem. 2018, 120, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.W.; Shin, N.R.; Lim, J.O.; Jung, T.Y.; Moon, C.; Kim, T.W.; Choi, J.; Shin, I.S.; Heo, J.D.; Kim, J.C. Silica dioxide nanoparticles aggravate airway inflammation in an asthmatic mouse model via NLRP3 inflammasome activation. Regul. Toxicol. Pharmacol. 2020, 112, 104618. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.O.; Ko, J.W.; Jung, T.Y.; Kim, W.I.; Pak, S.W.; Shin, I.S.; Yun, W.K.; Kim, H.C.; Heo, J.D.; Kim, J.C. Pulmonary inflammation caused by silica dioxide nanoparticles in mice via TXNIP/NLRP3 signaling pathway. Mol. Cell. Toxicol. 2020, 16, 245–252. [Google Scholar] [CrossRef]
- Zhou, W.; Shao, W.; Zhang, Y.; Liu, D.; Liu, M.; Jin, T. Glucagon-like peptide-1 receptor mediates the beneficial effect of liraglutide in an acute lung injury mouse model involving the thioredoxin-interacting protein. Am. J. Physiol. Endocrinol. Metab. 2020, 319, E568–E578. [Google Scholar] [CrossRef] [PubMed]
- Chihara, Y.; Iizumi, Y.; Horinaka, M.; Watanabe, M.; Goi, W.; Morita, M.; Nishimoto, E.; Sowa, Y.; Yamada, T.; Takayama, K.; et al. Histone deacetylase inhibitor OBP-801 and amrubicin synergistically inhibit the growth of squamous cell lung carcinoma by inducing mitochondrial ASK1-dependent apoptosis. Int. J. Oncol. 2020, 56, 848–856. [Google Scholar] [CrossRef]
- Deng, W.; Li, Y.; Jia, Y.; Tang, L.; He, Q.; Liu, D. Over-expression of thioredoxin-interacting protein promotes apoptosis of MIN6 cells via activating p38MAPK pathway. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi 2017, 33, 1323–1327. [Google Scholar]
- Izquierdo, J.L.; Almonacid, C.; González, Y.; Del Rio-Bermudez, C.; Ancochea, J.; Cárdenas, R.; Lumbreras, S.; Soriano, J.B. The impact of COVID-19 on patients with asthma. Eur. Respir. J. 2021, 57, 2003142. [Google Scholar] [CrossRef]
- Abdulnasser Harfoush, S.; Hannig, M.; Le, D.D.; Heck, S.; Leitner, M.; Omlor, A.J.; Tavernaro, I.; Kraegeloh, A.; Kautenburger, R.; Kickelbick, G.; et al. High-dose intranasal application of titanium dioxide nanoparticles induces the systemic uptakes and allergic airway inflammation in asthmatic mice. Respir. Res. 2020, 21, 168. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Chan, C.C.; Huang, W.C.; Kuo, M.L. Berberine inhibits pro-inflammatory cytokine-induced IL-6 and CCL11 production via modulation of STAT6 pathway in human bronchial epithelial cells. Int. J. Med. Sci. 2020, 17, 1464–1473. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Magaye, R.; Castranova, V.; Zhao, J. Titanium dioxide nanoparticles: A review of current toxicological data. Part. Fibre Toxicol. 2013, 10, 15. [Google Scholar] [CrossRef] [Green Version]
- Tanabe, T.; Rubin, B.K. Airway goblet cells secrete pro-inflammatory cytokines, chemokines, and growth factors. Chest 2016, 149, 714–720. [Google Scholar] [CrossRef]
- Manise, M.; Holtappels, G.; Van Crombruggen, K.; Schleich, F.; Bachert, C.; Louis, R. Sputum IgE and cytokines in asthma: Relationship with sputum cellular profile. PLoS ONE 2013, 8, e58388. [Google Scholar] [CrossRef]
- Mishra, V.; Baranwal, V.; Mishra, R.K.; Sharma, S.; Paul, B.; Pandey, A.C. Titanium dioxide nanoparticles augment allergic airway inflammation and Socs3 expression via NF-kappaB pathway in murine model of asthma. Biomaterials 2016, 92, 90–102. [Google Scholar] [CrossRef]
- Hsieh, C.C.; Papaconstantinou, J. Thioredoxin-ASK1 complex levels regulate ROS-mediated p38 MAPK pathway activity in livers of aged and long-lived Snell dwarf mice. FASEB J. 2006, 20, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Malone, C.F.; Emerson, C.; Ingraham, R.; Barbosa, W.; Guerra, S.; Yoon, H.; Liu, L.L.; Michor, F.; Haigis, M.; Macleod, K.F.; et al. mTOR and HDAC inhibitors converge on the TXNIP/thioredoxin pathway to cause catastrophic oxidative stress and regression of RAS-driven tumors. Cancer Discov. 2017, 7, 1450–1463. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.O.; Shin, N.R.; Seo, Y.S.; Nam, H.H.; Ko, J.W.; Jung, T.Y.; Lee, S.J.; Kim, H.J.; Cho, Y.K.; Kim, J.C.; et al. Silibinin attenuates silica dioxide nanoparticles-induced inflammation by suppressing TXNIP/MAPKs/AP-1 signaling. Cells 2020, 9, 678. [Google Scholar] [CrossRef] [Green Version]
- Shin, I.S.; Shin, N.R.; Park, J.W.; Jeon, C.M.; Hong, J.M.; Kwon, O.K.; Kim, J.S.; Lee, I.C.; Kim, J.C.; Oh, S.R.; et al. Melatonin attenuates neutrophil inflammation and mucus secretion in cigarette smoke-induced chronic obstructive pulmonary diseases via the suppression of Erk-Sp1 signaling. J. Pineal Res. 2015, 58, 50–60. [Google Scholar] [CrossRef]
- Okuda-Shimazaki, J.; Takaku, S.; Kanehira, K.; Sonezaki, S.; Taniguchi, A. Effects of titanium dioxide nanoparticle aggregate size on gene expression. Int. J. Mol. Sci. 2010, 11, 2383–2392. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BET Surface Area (m2/g) | ICP-MS (mg/g) | |||||||
|---|---|---|---|---|---|---|---|---|
| 40.45 | VC | TiO2NPs 5 | TiO2NPs 10 | TiO2NPs 20 | ||||
| 0.38 ± 0.032 | 2.88 ± 0.311 | 5.68 ± 0.597 | 6.69 ± 0.613 | |||||
| Single point Surface area (m2/g) | VC | OVA | OVA + TiO2NPs 5 | OVA + TiO2NPs 10 | OVA + TiO2NPs 20 | |||
| 39.38 | 0.34 ± 0.056 | 0.32 ± 0.042 | 2.53 ± 0.397 | 6.43 ± 0.617 | 7.94 ± 0.673 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, J.-O.; Lee, S.-J.; Kim, W.-I.; Pak, S.-W.; Moon, C.; Shin, I.-S.; Heo, J.-D.; Ko, J.-W.; Kim, J.-C. Titanium Dioxide Nanoparticles Exacerbate Allergic Airway Inflammation via TXNIP Upregulation in a Mouse Model of Asthma. Int. J. Mol. Sci. 2021, 22, 9924. https://doi.org/10.3390/ijms22189924
Lim J-O, Lee S-J, Kim W-I, Pak S-W, Moon C, Shin I-S, Heo J-D, Ko J-W, Kim J-C. Titanium Dioxide Nanoparticles Exacerbate Allergic Airway Inflammation via TXNIP Upregulation in a Mouse Model of Asthma. International Journal of Molecular Sciences. 2021; 22(18):9924. https://doi.org/10.3390/ijms22189924
Chicago/Turabian StyleLim, Je-Oh, Se-Jin Lee, Woong-Il Kim, So-Won Pak, Changjong Moon, In-Sik Shin, Jeong-Doo Heo, Je-Won Ko, and Jong-Choon Kim. 2021. "Titanium Dioxide Nanoparticles Exacerbate Allergic Airway Inflammation via TXNIP Upregulation in a Mouse Model of Asthma" International Journal of Molecular Sciences 22, no. 18: 9924. https://doi.org/10.3390/ijms22189924