
The Chemical Constituents from Fruits of Catalpa bignonioides Walt. and Their α-Glucosidase Inhibitory Activity and Insulin Secretion Effect


Abstract
:1. Introduction
2. Results and Discussion
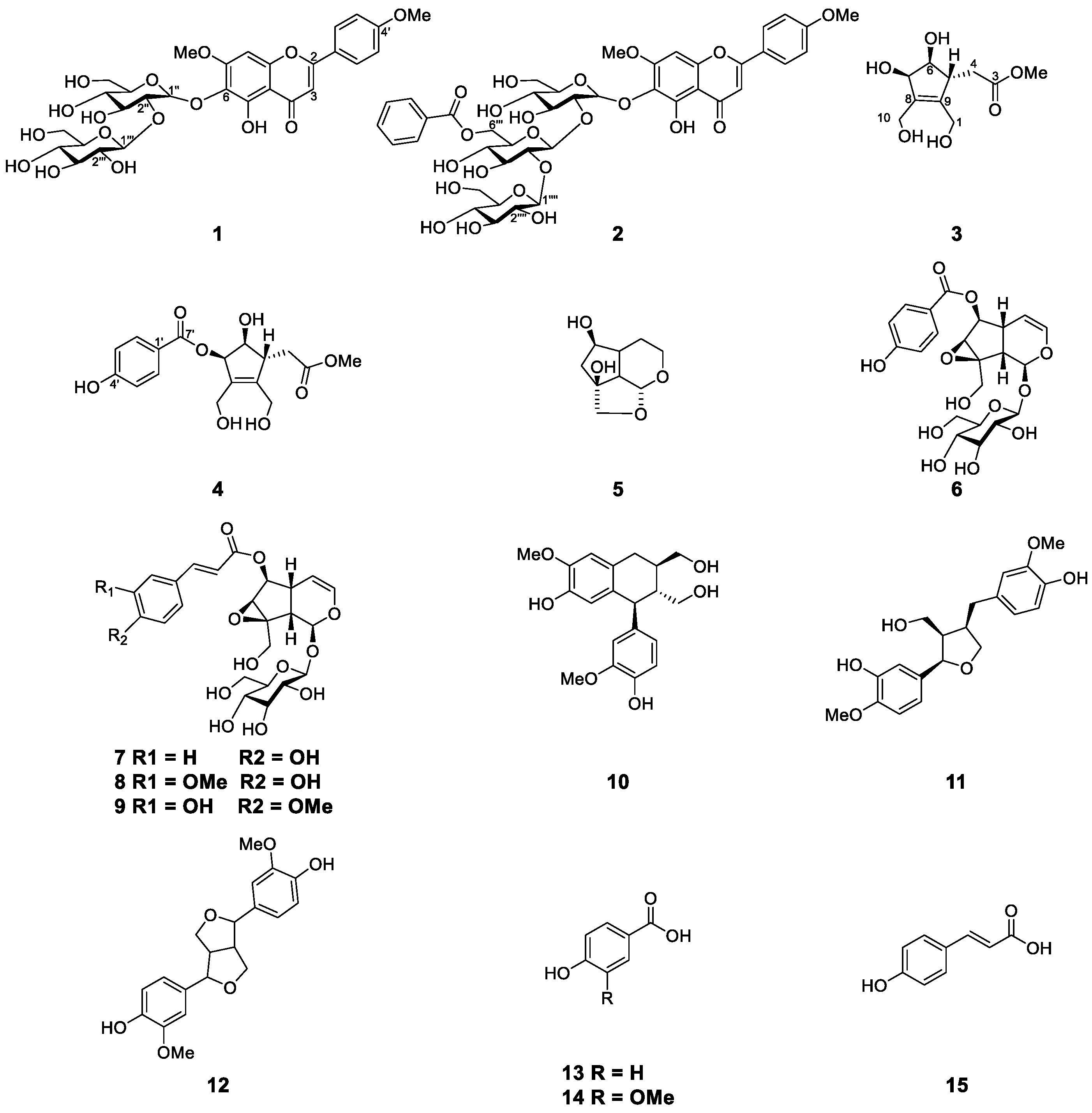
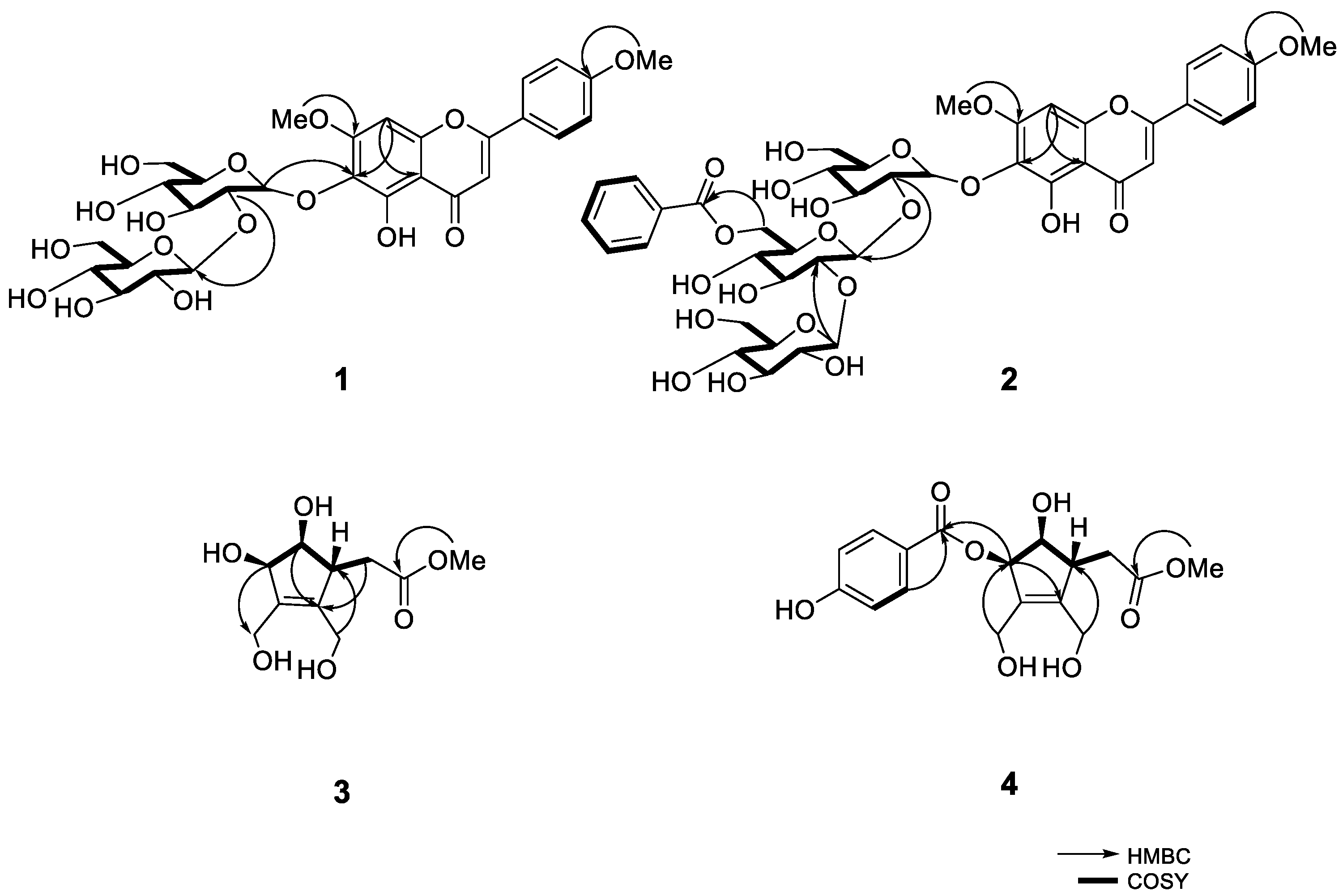
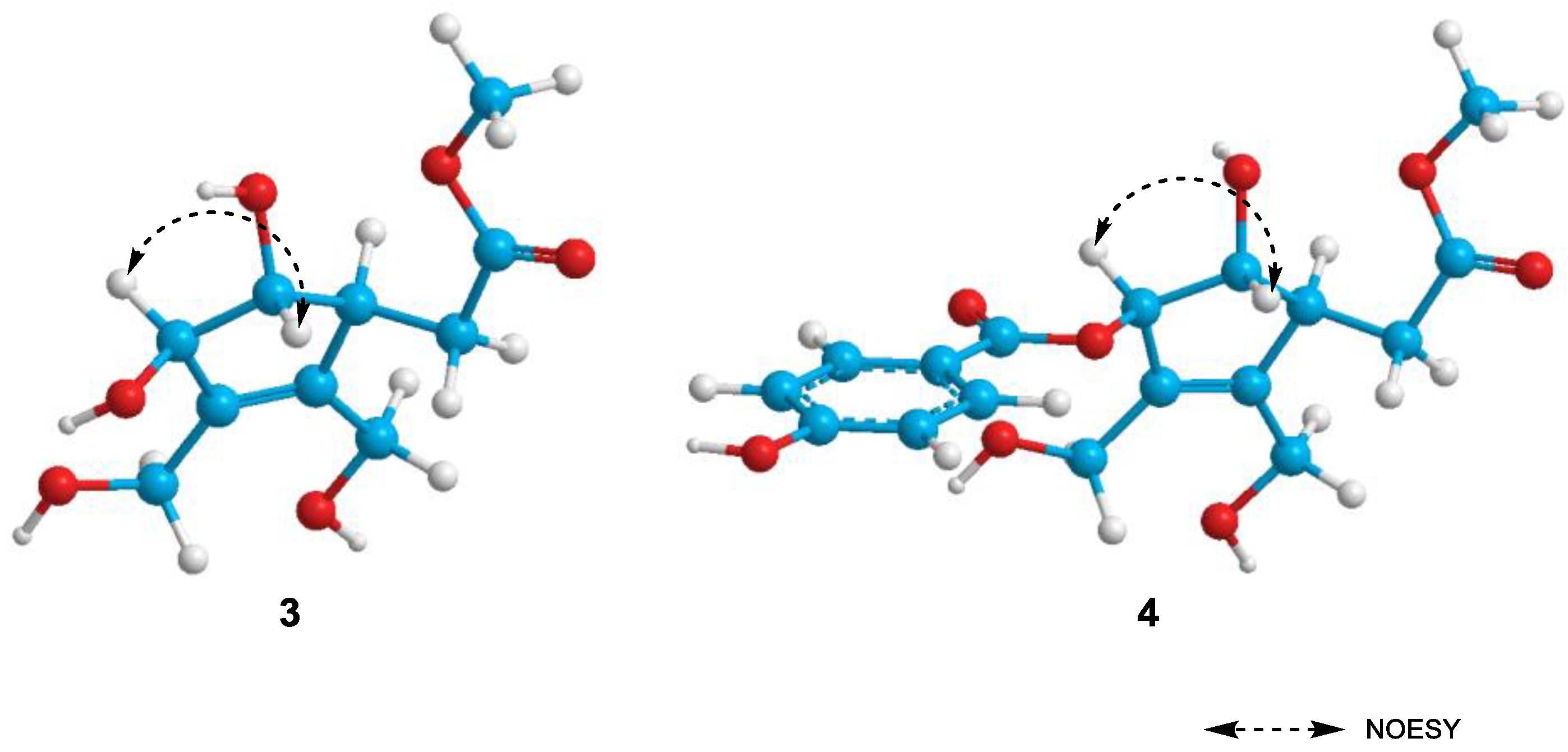
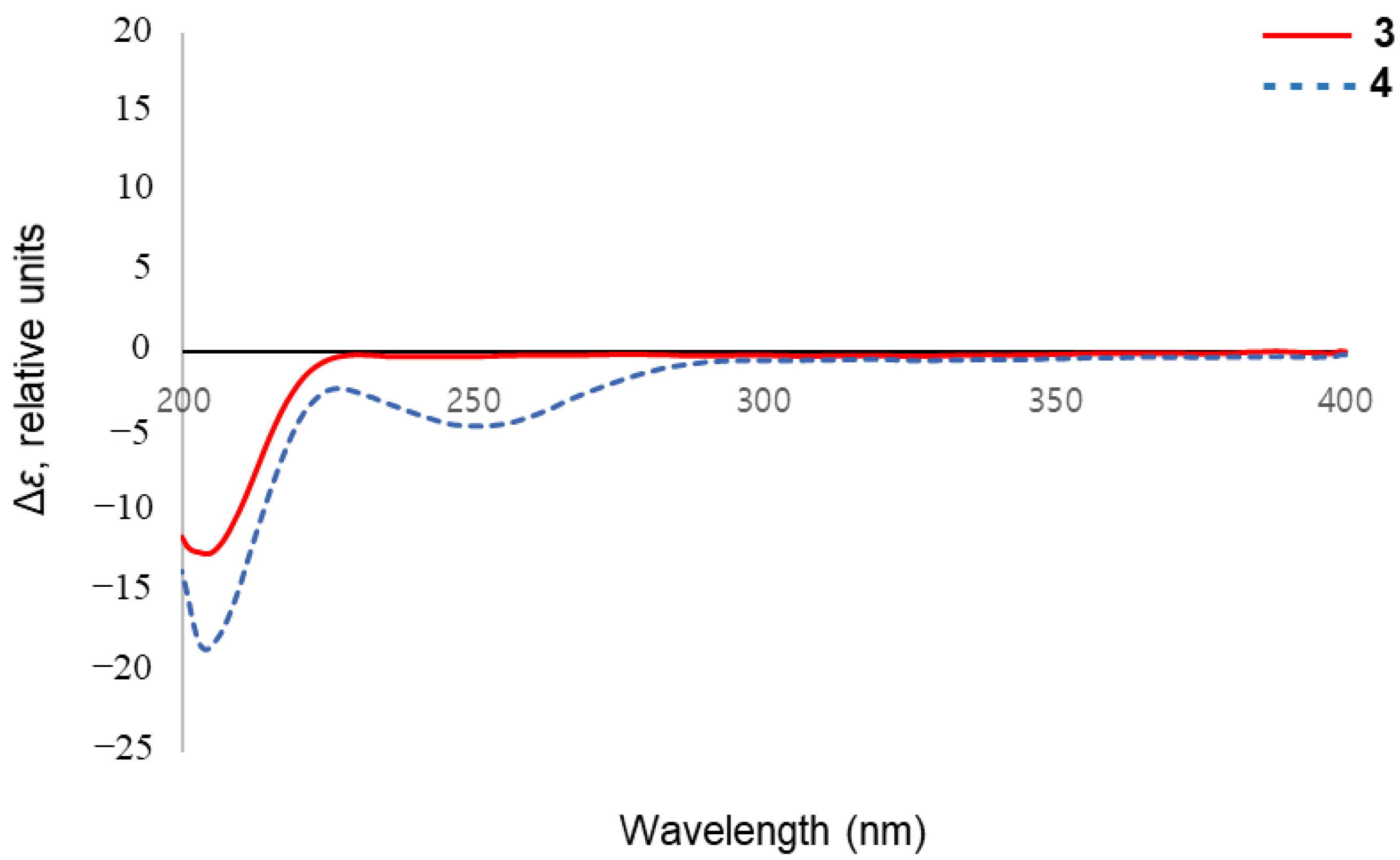
2.1. Structure Elucidation
2.2. α-Glucosidase Inhibitory Activity
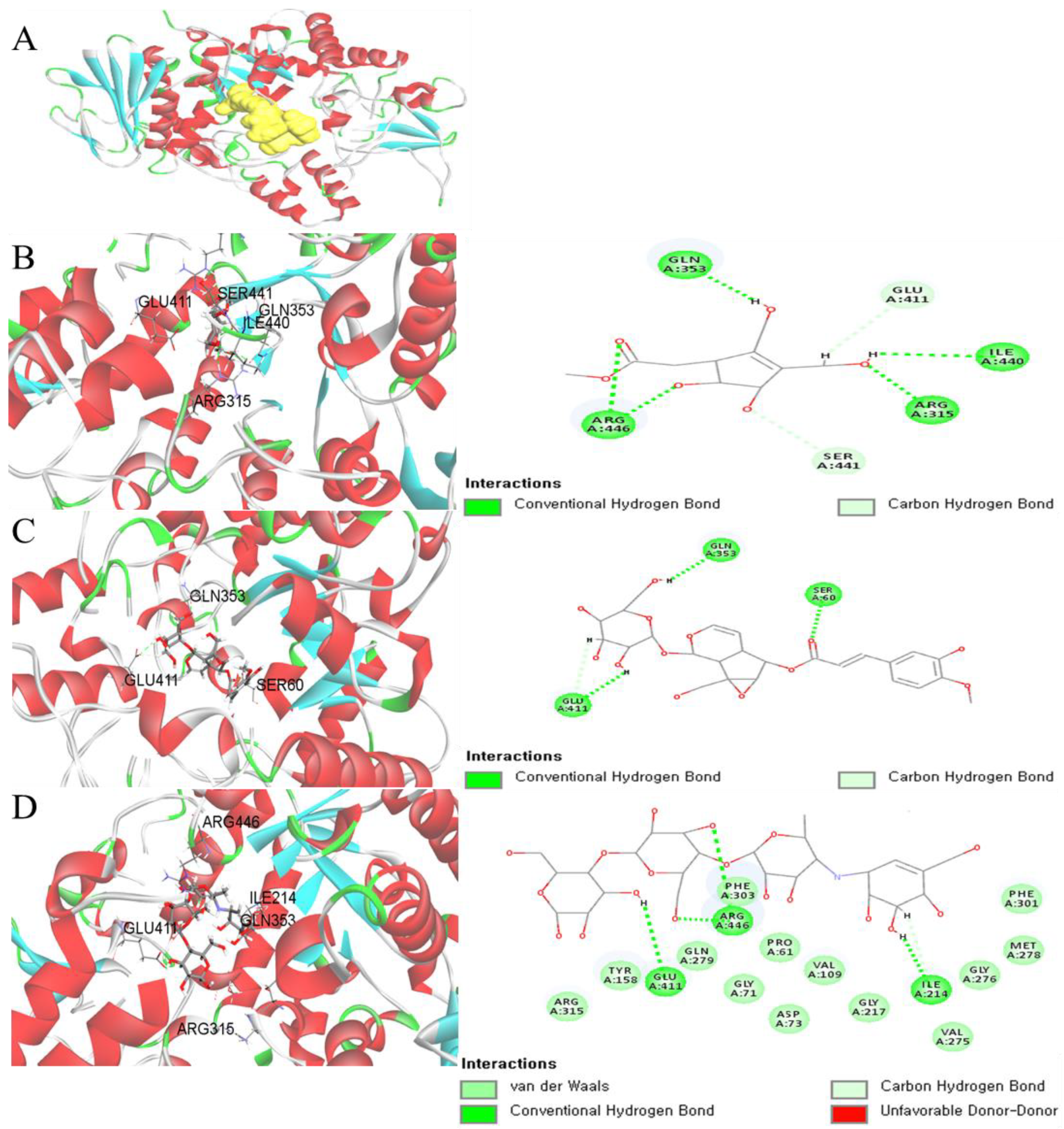
2.3. Virtual Screening of α-Glucosidase Inhibitors
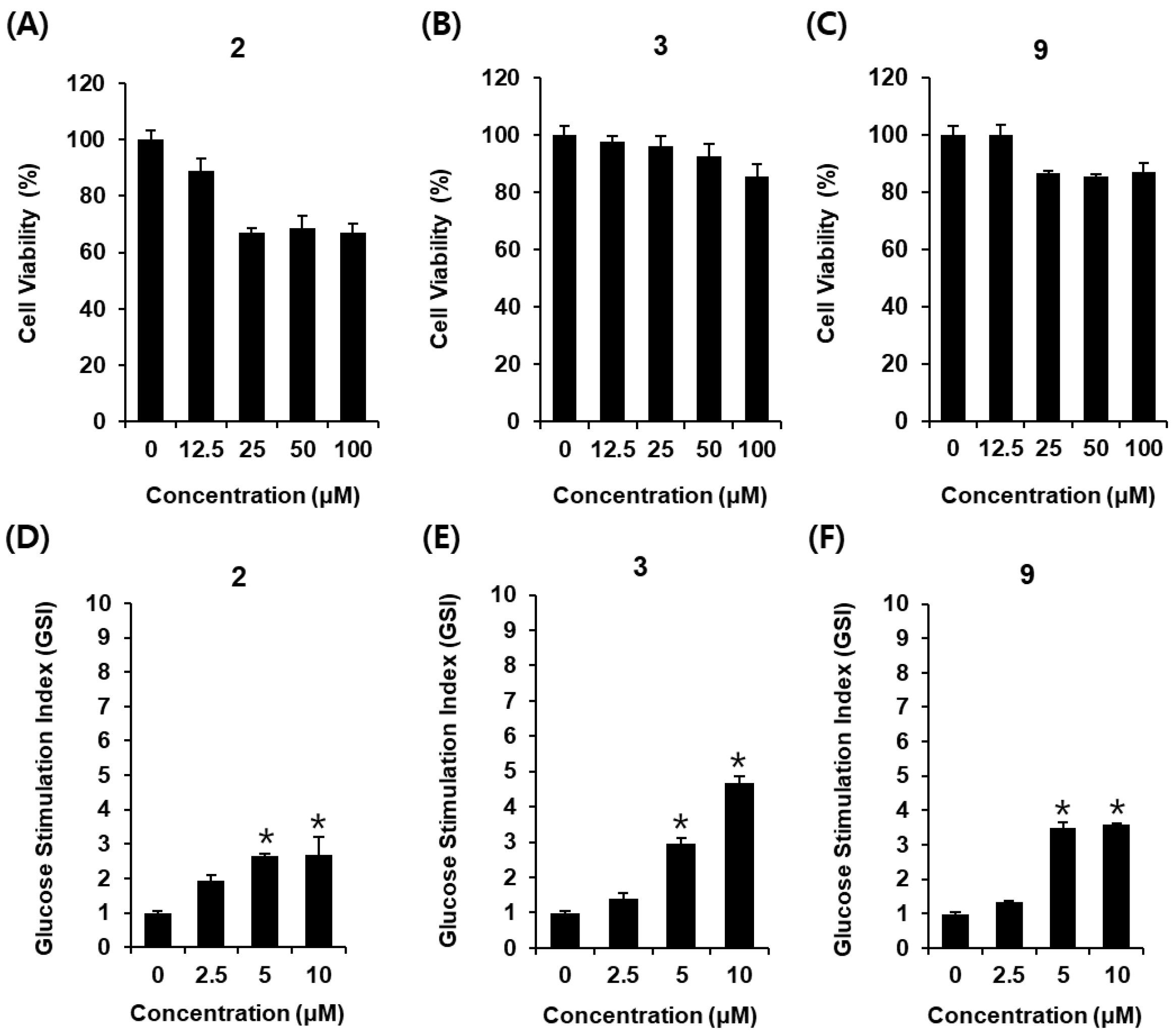
2.4. Glucose-Stimulated Insulin Secretion (GSIS) Effect
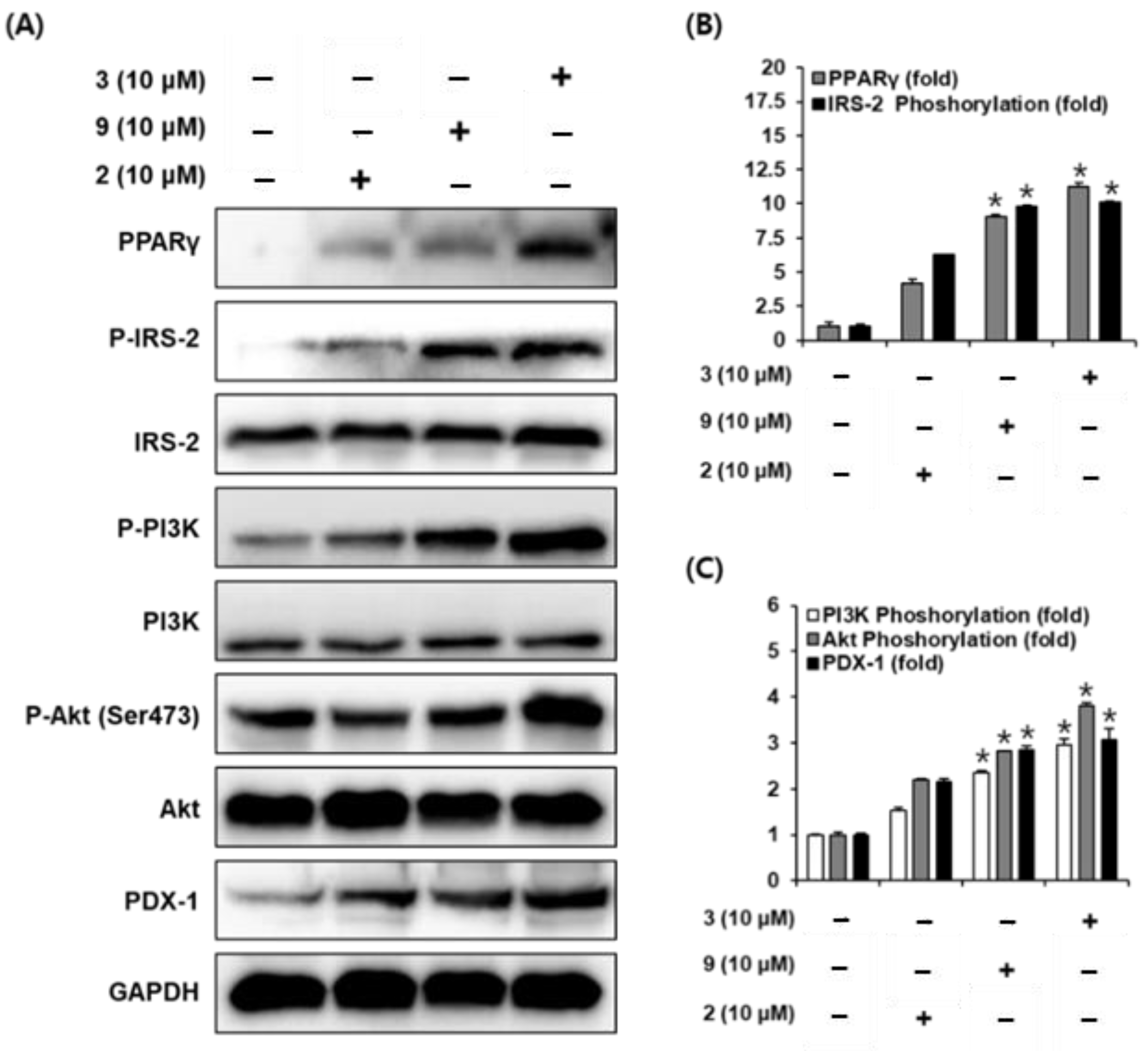
2.5. Protein Expression of PPARγ, P-IRS-2, IRS-2 (Ser731), P-PI3K, PI3K, P-Akt (Ser473), Akt, and PDX-1
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Plant Material
3.3. Isolation of Compounds 1–15
3.4. Assay of α-Glucosidase Activity
3.5. Structured-Based Virtual Screening
3.6. Cell Culture
3.7. Cell Viability Assay
3.8. GSIS Assay
3.9. Western Blot Analysis
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Lebovitz, H.E. Diagnosis, classification, and pathogenesis of diabetes mellitus. J. Clin. Psychiatry. 2001, 62, 5–9. [Google Scholar] [PubMed]
- Prabhakar, P.K.; Doble, M. Mechanism of action of natural products used in the treatment of diabetes mellitus. Chin. J. Integr. Med. 2011, 17, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Modak, M.; Dixit, P.; Londhe, J.; Ghaskadbi, S.; Devasagayam, T.P.A. Recent advances in Indian herbal drug research guest editor: Thomas Paul Asir Devasagayam Indian herbs and herbal drugs used for the treatment of diabetes. J. Clin. Biochem. 2007, 40, 163–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farag, M.; Ahmed, W.J.; Foud, I.; Mohammed, M.S. The Role of Medicinal Plants in the Treatment of Type-2 Diabetes. Adv. J. Pharm. Life Sci. Res. 2016, 4, 1–8. [Google Scholar]
- Munoz-Mingarro, D.; Acero, N.; Llinares, F.; Pozuelo, J.M.; Galan de Mera, A.; Vicenten, J.A.; Morales, L.; Alguacil, L.F.; Perez, C. Biological activity of extracts from Catalpa bignonioides Walt. (Bignoniaceae). J. Ethnopharmacol. 2003, 87, 163–167. [Google Scholar] [CrossRef]
- Dvorská, M.; Žemlička, M.; Muselík, J.; Karafiátová, J.; Suchý, V.J.F. Antioxidant activity of Catalpa bignonioides. Fitoterapia 2007, 78, 437–439. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Zhu, R.; Tian, Y.; Li, R.; Chen, B.; Zhang, H.; Xia, B.; Zhao, D.; Mo, F.; Zhang, D.J.M. Catalpol in Diabetes and its Complications: A Review of Pharmacology, Pharmacokinetics, and Safety. Molecules 2019, 24, 3302. [Google Scholar] [CrossRef] [Green Version]
- Hontecillas, R.; Diguardo, M.; Duran, E.; Orpi, M.; Bassaganya-Riera, J. Catalpic acid decreases abdominal fat deposition, improves glucose homeostasis and upregulates PPAR α expression in adipose tissue. Clin. Nutr. 2008, 27, 764–772. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Ono, I. Two new flavone glycosides from Catalpa ovata. Phytochemistry 1975, 14, 1654–1656. [Google Scholar] [CrossRef]
- Machida, K.; Ikeda, C.; Kakuda, R.; Yaoita, Y.; Kikuchi, M. Studies on the constituents of Catalpa species. V. Iridoids from Catalpae fructus. Nat. Med. 2001, 55, 61–63. [Google Scholar] [CrossRef]
- Kil, Y.S.; Kim, S.M.; Kang, U.; Chung, H.Y.; Seo, E.K. Peroxynitrite-Scavenging Glycosides from the Stem Bark of Catalpa ovata. J. Nat. Prod. 2017, 80, 2240–2251. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.H.; Kim, H.J.; Lee, K.H.; Kang, S.C.; Zee, O.P. Antioxidative iridoid glycosides and phenolic compounds from Veronica peregrina. Arch. Pharm. Res. 2009, 32, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.-X.; Xu, Y.; Bai, M.; Zhou, L.; Song, S.-J.; Wang, X.-B. Lignans from the seeds of Chinese hawthorn (Crataegus pinnatifida var. major NE Br.) against β-amyloid aggregation. Nat. Prod. Res. 2018, 32, 1706–1713. [Google Scholar] [CrossRef] [PubMed]
- Fedorova, T.E.; Fedorov, S.V.; Babkin, V.A. Oligolignans in the wood of Picea obovata Ledeb. Russ. J. Bioorg. Chem. 2016, 42, 712–715. [Google Scholar] [CrossRef]
- Hu, Q.W.; Chen, Y.Y.; Jiao, Q.Y.; Khan, A.; Shan, J.M.; Cao, G.D.; Li, F.; Zhang, C.; Lou, H.X. Polyphenolic compounds from Malus hupehensis and their free radical scavenging effects. Nat. Prod. Res. 2018, 32, 2152–2158. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.Q.; Xiao, J.; Fan, H.X.; Yu, Y.; He, R.R.; Feng, X.L.; Kurihara, H.; So, K.F.; Yao, X.S.; Gao, H. Polyphenols from wolfberry and their bioactivities. Food Chem. 2017, 214, 644–654. [Google Scholar] [CrossRef]
- Su, X.D.; Li, W.; Ma, J.Y.; Kim, Y.H. Chemical constituents from Epimedium koreanum Nakai and their chemotaxonomic significance. Nat. Prod. Res. 2018, 32, 2347–2351. [Google Scholar] [CrossRef]
- Kırmızıbekmez, H.; Bassarello, C.; Piacente, S.; Akaydın, G.; Çalış, İ. Flavonoid, phenylethanoid and iridoid glycosides from Globularia aphyllanthes. Z. Naturforsch. B. 2009, 64, 252–256. [Google Scholar] [CrossRef]
- Scott, A.; Wrixon, A. Stereochemistry of olefins—IX: Correlation of Mills’ and Brewster’s rules with the Cotton effects of cyclic olefins. Tetrahedron 1971, 27, 4787–4819. [Google Scholar] [CrossRef]
- Liu, Z.-X.; Liu, C.-T.; Liu, Q.-B.; Ren, J.; Li, L.-Z.; Huang, X.-X.; Wang, Z.-Z.; Song, S.-J. Iridoid glycosides from the flower buds of Lonicera japonica and their nitric oxide production and α-glucosidase inhibitory activities. J. Funct. Foods. 2015, 18, 512–519. [Google Scholar] [CrossRef]
- Kumar, S.; Narwal, S.; Kumar, V.; Prakash, O. α-glucosidase inhibitors from plants: A natural approach to treat diabetes. Pharmacogn. Rev. 2011, 5, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibano, M.; Kakutani, K.; Taniguchi, M.; Yasuda, M.; Baba, K. Antioxidant constituents in the dayflower (Commelina communis L.) and their α-glucosidase-inhibitory activity. J. Nat. Med. 2008, 62, 349. [Google Scholar] [CrossRef] [PubMed]
- Hussain, N.; Hameed, A.; Ahmad, M.S.; Yousuf, S.; Ali, Z.; Rahman, M.H.; Choudhary, M.I. New iridoids from Lyonia ovalifolia and their anti-hyperglycemic effects in mice pancreatic islets. Fitoterapia 2018, 131, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Kappel, V.D.; Frederico, M.J.; Postal, B.G.; Mendes, C.P.; Cazarolli, L.H.; Silva, F.R.M.B. The role of calcium in intracellular pathways of rutin in rat pancreatic islets: Potential insulin secretagogue effect. Eur. J. Pharmacol. 2013, 702, 264–268. [Google Scholar] [CrossRef] [Green Version]
- Gupta, D.; Kono, T.; Evans-Molina, C. The role of peroxisome proliferator-activated receptor γ in pancreatic β cell function and survival: Therapeutic implications for the treatment of type 2 diabetes mellitus. Diabetes Obes. Metab. 2010, 12, 1036–1047. [Google Scholar] [CrossRef] [Green Version]
- Moibi, J.A.; Gupta, D.; Jetton, T.L.; Peshavaria, M.; Desai, R.; Leahy, J.L. Peroxisome Proliferator–Activated Receptor-γ Regulates Expression of PDX-1 and NKX6. 1 in INS-1 Cells. Diabetes 2007, 56, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Gupta, D.; Jetton, T.L.; Mortensen, R.M.; Duan, S.Z.; Peshavaria, M.; Leahy, J.L. In vivo and in vitro studies of a functional peroxisome proliferator-activated receptor γ response element in the mouse pdx-1 promoter. J. Biol. Chem. 2008, 283, 32462–32470. [Google Scholar] [CrossRef] [Green Version]
- Roncero, I.; Alvarez, E.; Acosta, C.; Sanz, C.; Barrio, P.; Hurtado-Carneiro, V.; Burks, D.; Blazquez, E. Insulin-receptor substrate-2 (irs-2) is required for maintaining glucokinase and glucokinase regulatory protein expression in mouse liver. PLoS ONE 2013, 8, e58797. [Google Scholar] [CrossRef]
- Liu, S.; Li, X.; Wu, Y.; Duan, R.; Zhang, J.; Du, F.; Zhang, Q.; Li, Y.; Li, N. Effects of vaspin on pancreatic β cell secretion via PI3K/Akt and NF-κB signaling pathways. PLoS ONE 2017, 12, e0189722. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Liao, G.; Xiang, C.; Yang, X.; Cheng, X.; Ou, Y. Effects of phycocyanin on INS-1 pancreatic β-cell mediated by PI3K/Akt/FoxO1 signaling pathway. J. Biol. Macromol. 2016, 83, 185–194. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 | 2 | ||||
|---|---|---|---|---|---|
| Pos. | δCa,b | δHa,d (J in Hz) | Pos. | δCa,b | δHa,c (J in Hz) |
| 1 | 1 | - | |||
| 2 | 163.5 | - | 2 | 164.1 | - |
| 3 | 103.4 | 6.90 (s) | 3 | 102.9 | 6.53 (s) |
| 4 | 182.2 | - | 4 | 182.5 | - |
| 5 | 151.0 | - | 5 | 152.8 | - |
| 6 | 127.9 | - | 6 | 128.0 | - |
| 7 | 158.3 | - | 7 | 158.4 | - |
| 8 | 91.9 | 6.92 (s) | 8 | 90.9 | 6.43 (s) |
| 9 | 152.5 | - | 9 | 153.1 | - |
| 10 | 104.9 | - | 10 | 105.0 | - |
| 1’ | 122.8 | - | 1’ | 123.0 | - |
| 2’, 6’ | 128.4 | 8.04 (d, 8.9) | 2’, 6’ | 127.8 | 7.90 (d, 8.4) |
| 3’, 5’ | 114.6 | 7.09 (d, 9.0) | 3’, 5’ | 114.2 | 7.08 (d, 8.4) |
| 4’ | 162.4 | - | 4’ | 162.9 | - |
| 1’’ | 99.6 | 5.30 (d, 6.5) | 1’’ | 100.4 | 5.28 (d, 7.0) |
| 2’’ | 81.3 | 3.63 * | 2’’ | 85.0 | 3.62 * |
| 3’’ | 76.3 | 3.45 * | 3’’ | 75.7 | 3.65 * |
| 4’’ | 69.9 | 3.05 * | 4’’ | 68.8 | 3.50 * |
| 5’’ | 77.0 | 3.15 * | 5’’ | 76.1 | 3.20 * |
| 6’’ | 60.9 | 3.54 * | 6’’ | 60.9 | 3.67 * |
| 1’’’ | 102.9 | 4.57 (d, 7.9) | 1’’’ | 102.9 | 4.74 (d, 7.6) |
| 2’’’ | 74.1 | 2.98 * | 2’’’ | 83.1 | 3.56 * |
| 3’’’ | 76.4 | 3.12 * | 3’’’ | 76.6 | 3.64 * |
| 4’’’ | 69.2 | 3.33 * | 4’’’ | 69.6 | 3.34 * |
| 5’’’ | 76.9 | 3.05 * | 5’’’ | 76.9 | 3.20 * |
| 6’’’ | 60.8 | 3.40 * | 6’’’ | 64.0 | 4.33 (d, 3.8) |
| 1’’’’ | 104.4 | 4.71 (d, 7.7) | |||
| 2’’’’ | 74.4 | 3.28 * | |||
| 3’’’’ | 76.3 | 3.37 * | |||
| 4’’’’ | 69.9 | 3.37 * | |||
| 5’’’’ | 77.1 | 3.35 * | |||
| 6’’’’ | 60.9 | 3.62 * | |||
| 1’’’’’ | 129.2 | - | |||
| 2’’’’’, 6’’’’’ | 128.7 | 7.90 (m) | |||
| 3’’’’’, 5’’’’’ | 127.5 | 7.13 (t, 7.4) | |||
| 4’’’’’ | 132.4 | 7.29 (t, 7.4) | |||
| 7’’’’’ | 166.3 | - | |||
| 4’-OMe | 55.6 | 3.82 (s) | 4’-OMe | 54.6 | 3.88 (s) |
| 7-OMe | 56.6 | 3.87 (s) | 7-OMe | 55.4 | 3.83 (s) |
| 3 | 4 | |||
|---|---|---|---|---|
| Pos. | δCa,b | δHa,c (J in Hz) | δCa,b | δHa,c (J in Hz) |
| 1 | 56.6 | 4.27 * 4.09 * | 56.6 | 4.33 * 4.11 * |
| 3 | 174.9 | - | 174.7 | - |
| 4 | 35.9 | 2.77 (dd, 15.5, 5.5) 2.48 (dd, 15.5, 8.2) | 35.8 | 2.78 (dd, 15.6, 5.7) 2.52 (dd, 15.6, 7.8) |
| 5 | 49.2 | 3.17 (m) | 49.9 | 3.24 (m) |
| 6 | 76.5 | 3.95 (t, 5.0) | 75.8 | 4.17 * |
| 7 | 75.5 | 4.56 (d, 5.4) | 78.3 | 5.87 (d, 5.8) |
| 8 | 139.6 | - | 136.7 | - |
| 9 | 142.3 | - | 145.3 | - |
| 10 | 57.2 | 4.29 * 4.15 * | 57.2 | 4.20 * 4.17 * |
| 1’ | 122.5 | - | ||
| 2’, 6’ | 133.0 | 7.94 (d, 3.2) | ||
| 3’, 5’ | 116.0 | 6.83 (d, 1.9) | ||
| 4’ | 163.4 | - | ||
| 7’ | 168.1 | - | ||
| 3-OMe | 52.1 | 3.75 (s) | 52.2 | 3.58 (s) |
| Compound | α-Glucosidase (units/L) |
|---|---|
| Acarbose | 0.41 ± 0.25 |
| 1 | 0.82 ± 0.59 |
| 2 | 0.57 ± 0.64 |
| 3 | 0.43 ± 0.19 |
| 4 | 0.77 ± 0.11 |
| 5 | 1.31 ± 0.45 |
| 6 | 1.21 ± 0.15 |
| 7 | 1.41 ± 0.28 |
| 8 | 2.44 ± 0.32 |
| 9 | 0.41 ± 1.09 |
| 10 | 1.10 ± 1.03 |
| 11 | 1.08 ± 0.52 |
| 12 | 6.35 ± 1.70 |
| 13 | 0.67 ± 0.60 |
| 14 | 0.75 ± 0.35 |
| 15 | 0.67 ± 0.58 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, Y.; Lee, D.; Park, S.; Kim, S.H.; Kang, K.S. The Chemical Constituents from Fruits of Catalpa bignonioides Walt. and Their α-Glucosidase Inhibitory Activity and Insulin Secretion Effect. Molecules 2021, 26, 362. https://doi.org/10.3390/molecules26020362
Oh Y, Lee D, Park S, Kim SH, Kang KS. The Chemical Constituents from Fruits of Catalpa bignonioides Walt. and Their α-Glucosidase Inhibitory Activity and Insulin Secretion Effect. Molecules. 2021; 26(2):362. https://doi.org/10.3390/molecules26020362
Chicago/Turabian StyleOh, Youngse, Dahae Lee, SeonJu Park, Seung Hyun Kim, and Ki Sung Kang. 2021. "The Chemical Constituents from Fruits of Catalpa bignonioides Walt. and Their α-Glucosidase Inhibitory Activity and Insulin Secretion Effect" Molecules 26, no. 2: 362. https://doi.org/10.3390/molecules26020362