
Decursinol Angelate Mitigates Sepsis Induced by Methicillin-Resistant Staphylococcus aureus Infection by Modulating the Inflammatory Responses of Macrophages

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
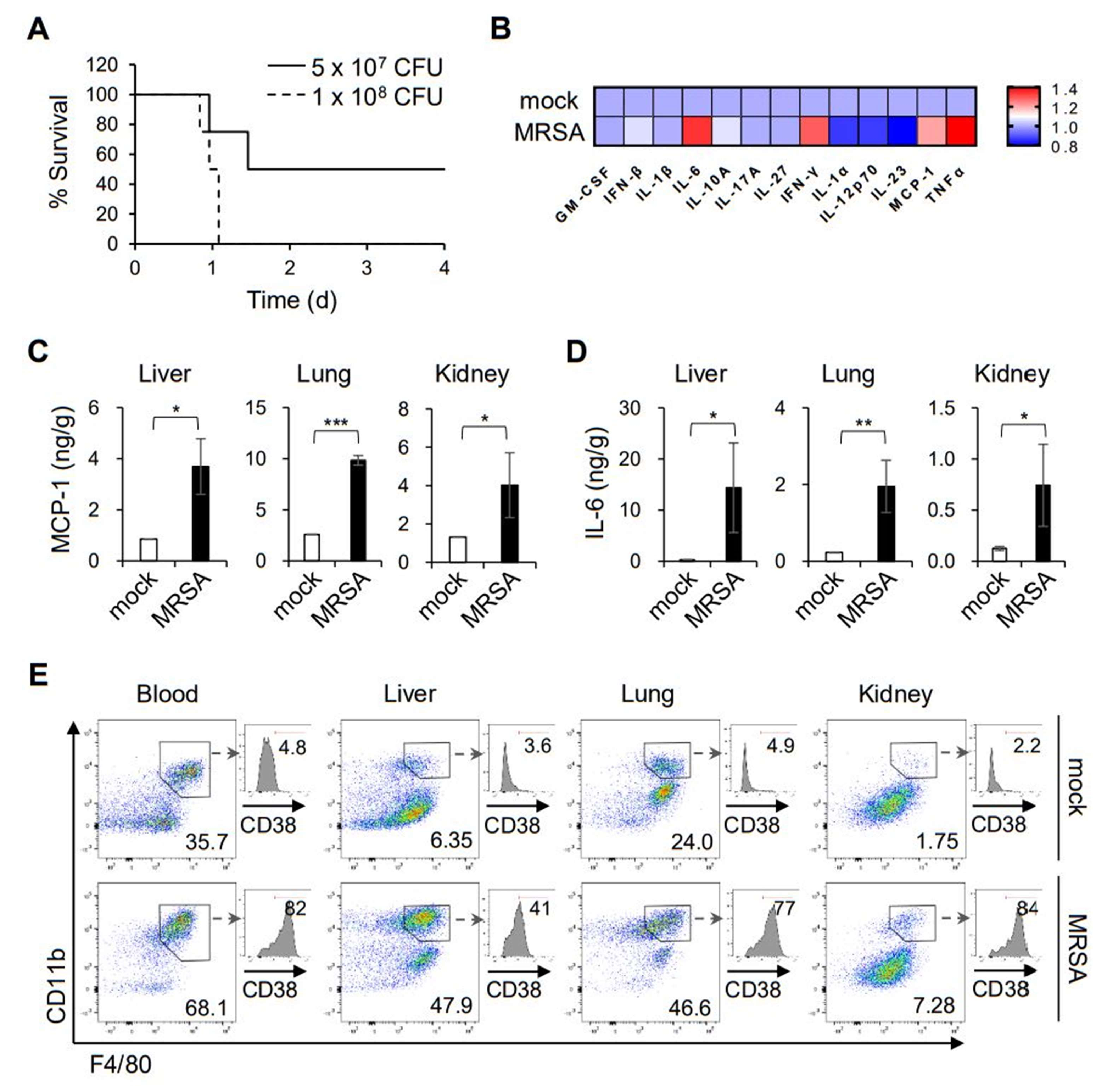
2.1. MRSA Infection Activates Macrophages Associated with the Pro-Inflammation Phenotype
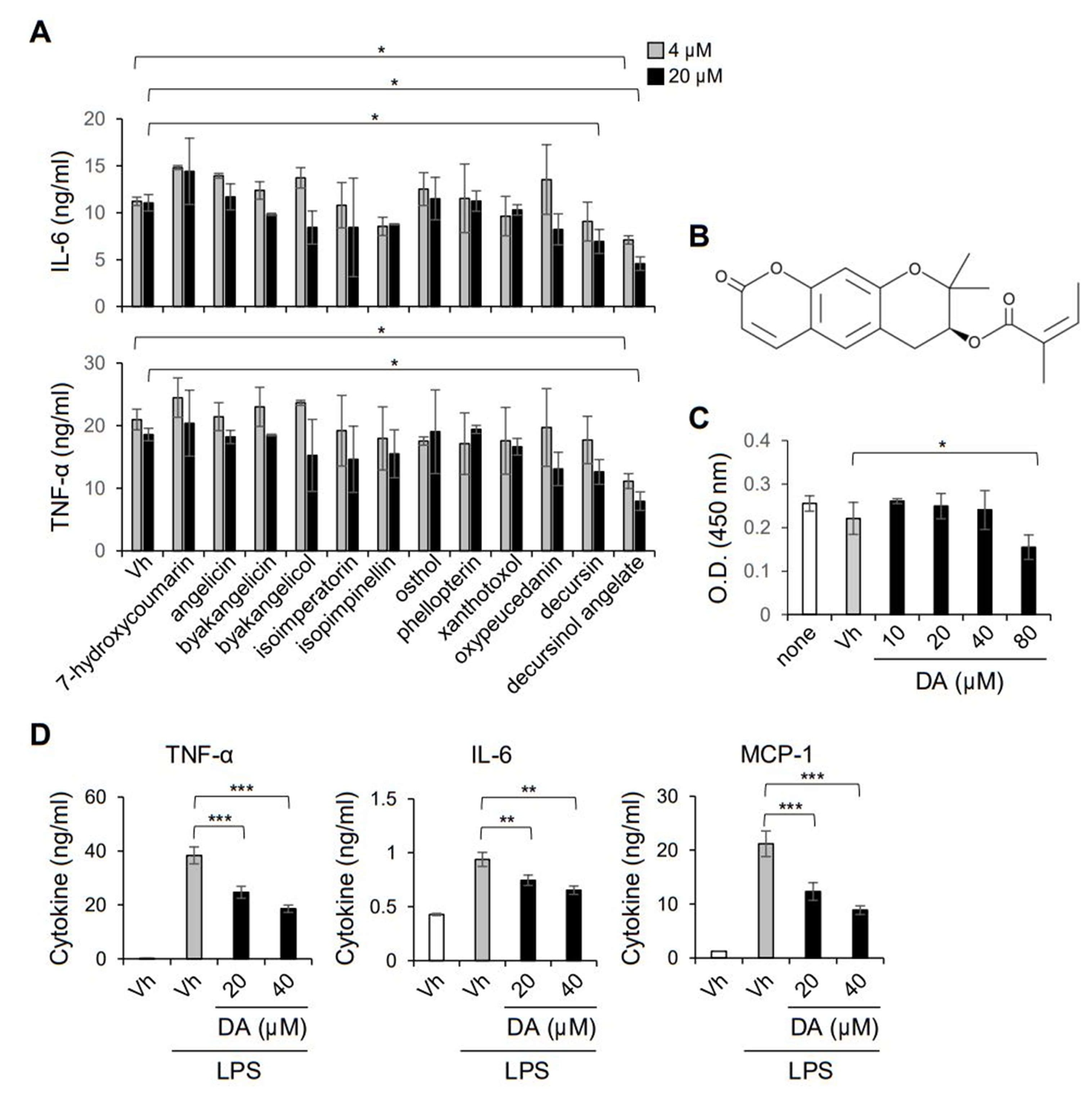
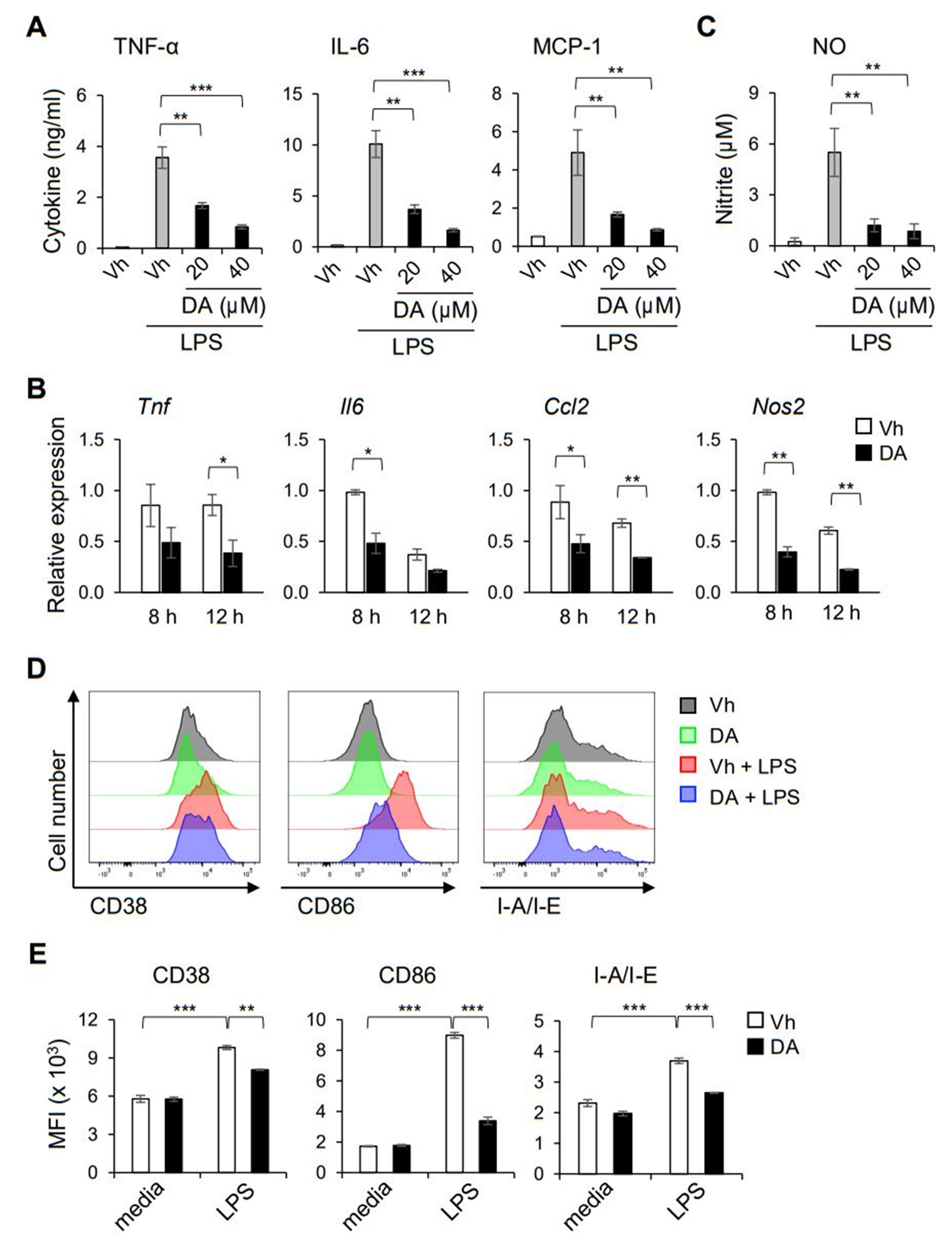
2.2. DA Inhibits LPS-Induced Production of Pro-Inflammatory Cytokines in Macrophages
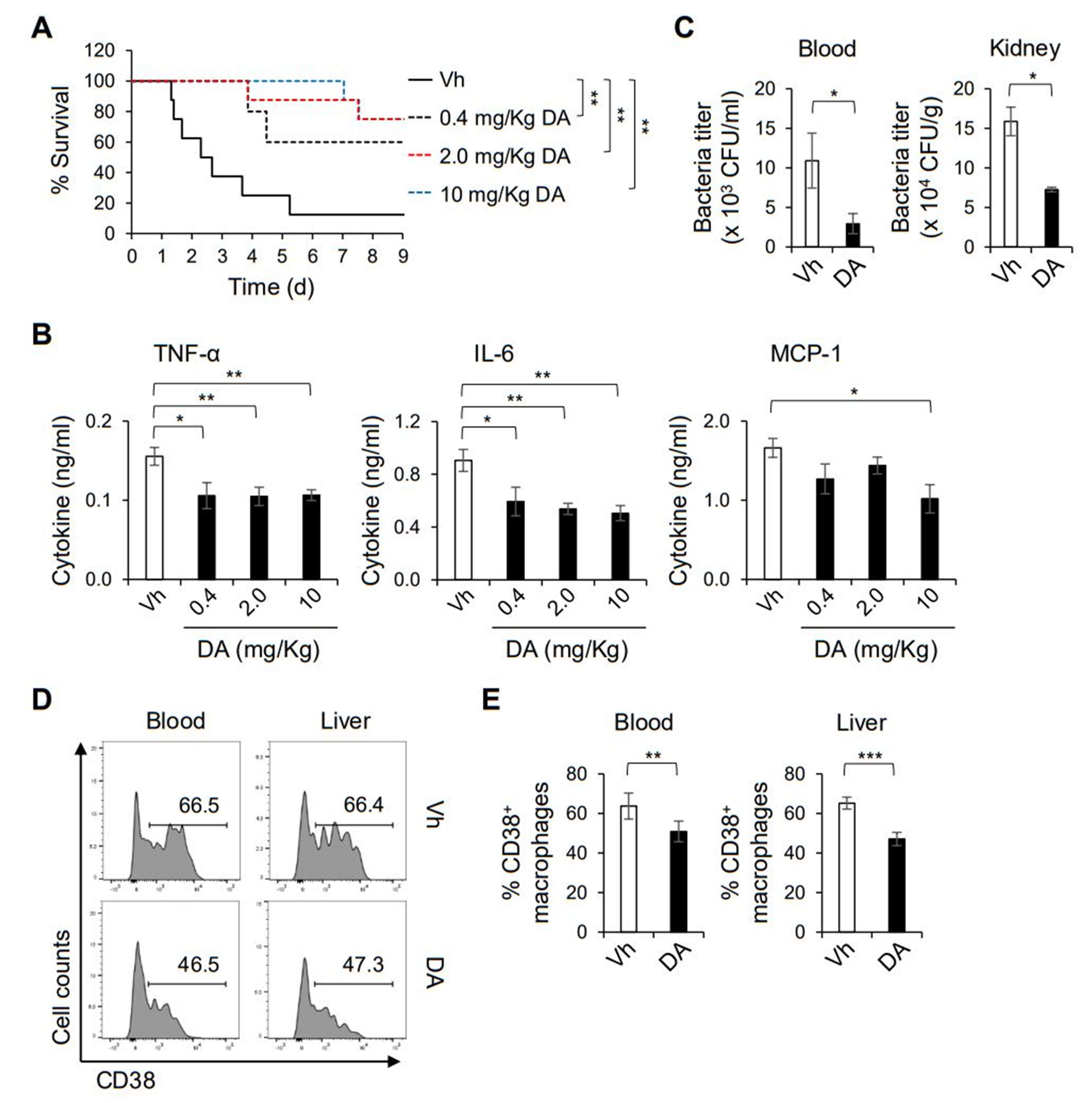
2.3. DA Alleviates Septic Symptoms Induced by MRSA Infection
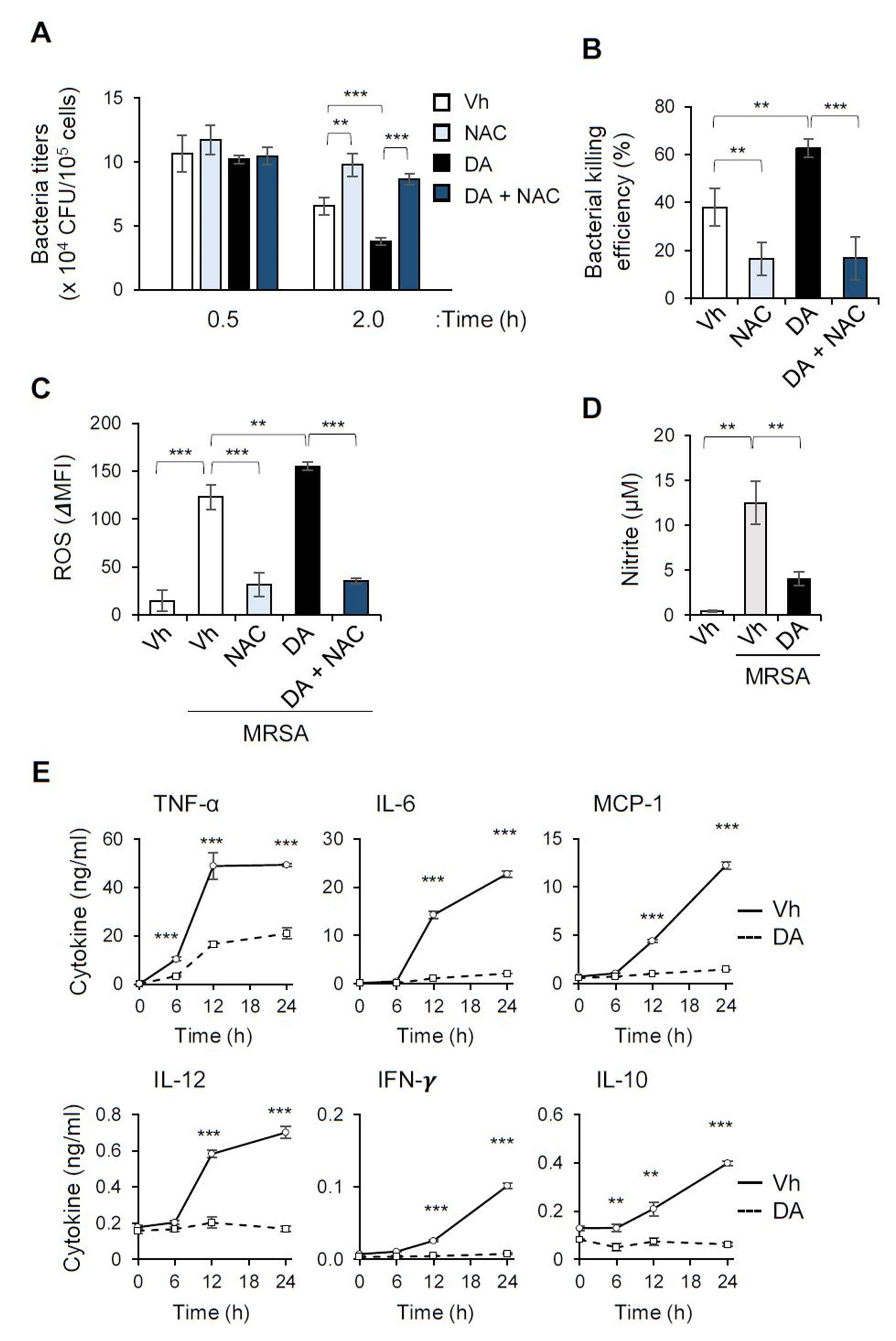
2.4. DA Enhances the Bacterial Killing Ability of Macrophages While Suppressing the Induction of Pro-Inflammatory Cytokines in Response to MRSA
2.5. DA Modulates the Functional Activation of Macrophages
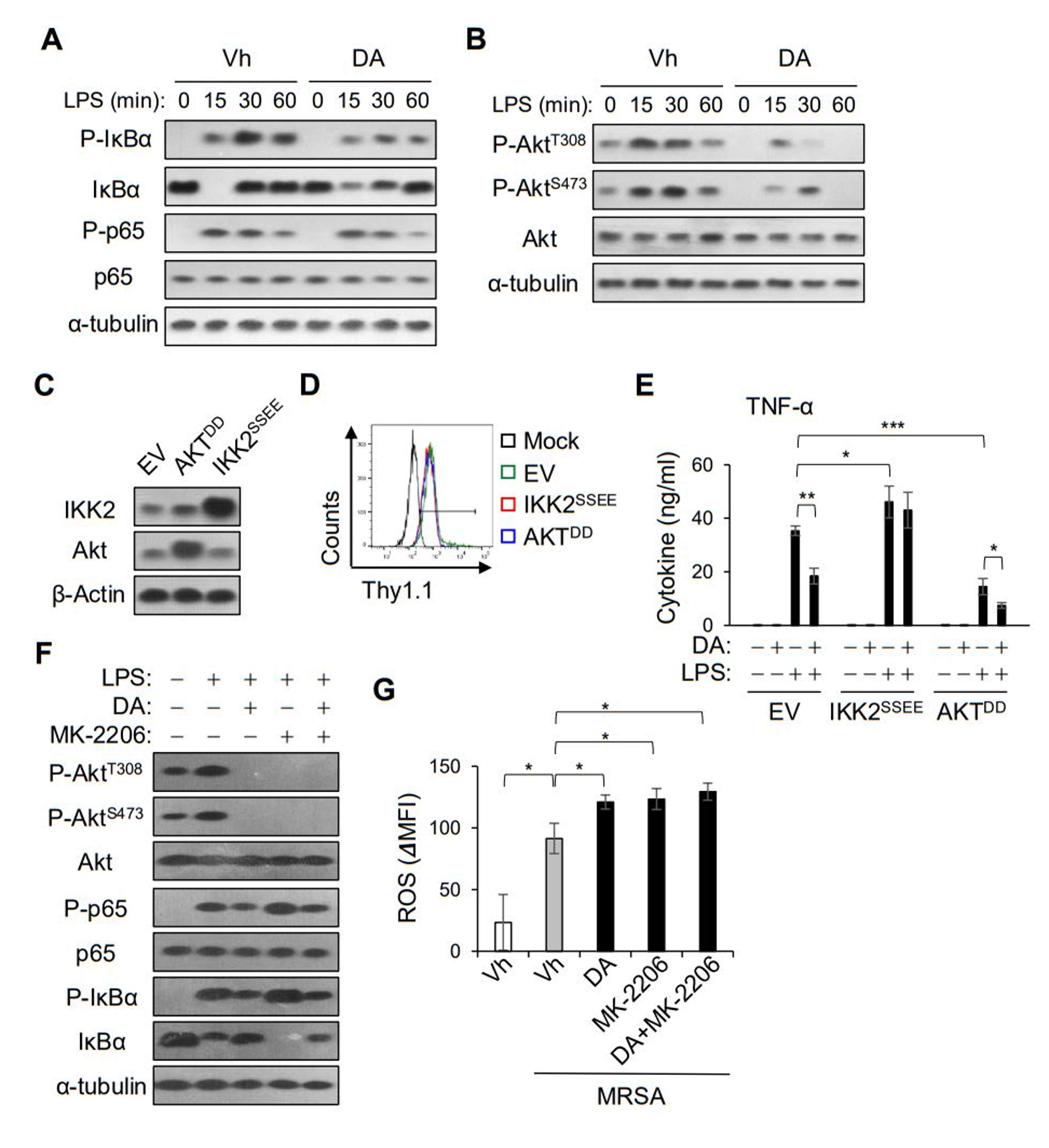
2.6. DA Suppressed the NF-κB Pathway Independently of Akt Inhibition
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Bacteria Culture and Colony Formation Assay
4.3. Mice
4.4. Macrophages
4.5. Cell Viability Assay
4.6. Flow Cytometry and Cytokine Measurement
4.7. Phagocytosis and Bacteria-Killing Assay
4.8. Measurement of Reactive Oxygen Species and Nitric Oxide
4.9. Quantitative Real-Time PCR
4.10. Western Blotting
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Thapa, B.; Lee, K. Metabolic influence on macrophage polarization and pathogenesis. BMB Rep. 2019, 52, 360–372. [Google Scholar] [CrossRef] [PubMed]
- Kaiko, G.E.; Horvat, J.C.; Beagley, K.W.; Hansbro, P.M. Immunological decision-making: How does the immune system decide to mount a helper T-cell response? Immunology 2008, 123, 326–338. [Google Scholar] [CrossRef]
- Gordon, S. Alternative activation of macrophages. Nat. Rev. Immunol. 2003, 3, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A.; Vannella, K.M. Macrophages in Tissue Repair, Regeneration, and Fibrosis. Immunity 2016, 44, 450–462. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, B.E.; Martinez-Aguilar, G.; Hulten, K.G.; Hammerman, W.A.; Coss-Bu, J.; Avalos-Mishaan, A.; Mason, E.O., Jr.; Kaplan, S.L. Severe Staphylococcal sepsis in adolescents in the era of community-acquired methicillin-resistant Staphylococcus aureus. Pediatrics 2005, 115, 642–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeLeo, F.R.; Chambers, H.F. Reemergence of antibiotic-resistant Staphylococcus aureus in the genomics era. J. Clin. Investig. 2009, 119, 2464–2474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Li, L.; Jiang, C.; Xing, C.; Kim, S.H.; Lu, J. Anti-cancer and other bioactivities of Korean Angelica gigas Nakai (AGN) and its major pyranocoumarin compounds. Anticancer Agents Med. Chem. 2012, 12, 1239–1254. [Google Scholar] [CrossRef]
- Thapa, B.; Pak, S.; Kwon, H.J.; Lee, K. Decursinol Angelate Ameliorates Dextran Sodium Sulfate-Induced Colitis by Modulating Type 17 Helper T Cell Responses. Biomol. Ther. 2019, 27, 466–473. [Google Scholar] [CrossRef]
- Song, G.Y.; Lee, J.H.; Cho, M.; Park, B.S.; Kim, D.E.; Oh, S. Decursin suppresses human androgen-independent PC3 prostate cancer cell proliferation by promoting the degradation of beta-catenin. Mol. Pharmacol. 2007, 72, 1599–1606. [Google Scholar] [CrossRef] [Green Version]
- Shehzad, A.; Islam, S.U.; Ahn, E.M.; Lee, Y.M.; Lee, Y.S. Decursinol angelate inhibits PGE2-induced survival of the human leukemia HL-60 cell line via regulation of the EP2 receptor and NFkappaB pathway. Cancer Biol. Ther. 2016, 17, 985–993. [Google Scholar] [CrossRef] [Green Version]
- Yang, E.J.; Song, G.Y.; Lee, J.S.; Yun, C.Y.; Kim, I.S. A novel (S)-(+)-decursin derivative, (S)-(+)-3-(3,4-dihydroxy-phenyl)-acrylic acid 2,2-dimethyl-8-oxo-3,4-dihydro-2H,8H-pyrano[3,2-g]chromen-3-yl-ester, inhibits ovalbumin-induced lung inflammation in a mouse model of asthma. Biol. Pharm. Bull. 2009, 32, 444–449. [Google Scholar] [CrossRef] [Green Version]
- Shehzad, A.; Parveen, S.; Qureshi, M.; Subhan, F.; Lee, Y.S. Decursin and decursinol angelate: Molecular mechanism and therapeutic potential in inflammatory diseases. Inflamm. Res. 2018, 67, 209–218. [Google Scholar] [CrossRef]
- Kim, J.H.; Jeong, J.H.; Jeon, S.T.; Kim, H.; Ock, J.; Suk, K.; Kim, S.I.; Song, K.S.; Lee, W.H. Decursin inhibits induction of inflammatory mediators by blocking nuclear factor-kappaB activation in macrophages. Mol. Pharmacol. 2006, 69, 1783–1790. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.U.; Lee, J.H.; Shehzad, A.; Ahn, E.M.; Lee, Y.M.; Lee, Y.S. Decursinol Angelate Inhibits LPS-Induced Macrophage Polarization through Modulation of the NFkappaB and MAPK Signaling Pathways. Molecules 2018, 23, 1880. [Google Scholar] [CrossRef] [Green Version]
- Sastalla, I.; Monack, D.M.; Kubatzky, K.F. Editorial: Bacterial Exotoxins: How Bacteria Fight the Immune System. Front. Immunol. 2016, 7, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jablonski, K.A.; Amici, S.A.; Webb, L.M.; Ruiz-Rosado, J.d.D.; Popovich, P.G.; Partida-Sanchez, S.; Guerau-de-Arellano, M. Novel Markers to Delineate Murine M1 and M2 Macrophages. PLoS ONE 2015, 10, e0145342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joo, S.S.; Park, D.; Shin, S.; Jeon, J.H.; Kim, T.K.; Choi, Y.J.; Lee, S.H.; Kim, J.S.; Park, S.K.; Hwang, B.Y.; et al. Anti-allergic effects and mechanisms of action of the ethanolic extract of Angelica gigas in dinitrofluorobenzene-induced inflammation models. Environ. Toxicol. Pharmacol. 2010, 30, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Surewaard, B.G.; Deniset, J.F.; Zemp, F.J.; Amrein, M.; Otto, M.; Conly, J.; Omri, A.; Yates, R.M.; Kubes, P. Identification and treatment of the Staphylococcus aureus reservoir in vivo. J. Exp. Med. 2016, 213, 1141–1151. [Google Scholar] [CrossRef] [Green Version]
- Uribe-Querol, E.; Rosales, C. Control of Phagocytosis by Microbial Pathogens. Front. Immunol. 2017, 8, 1368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goswami, M.; Sharma, D.; Khan, N.M.; Checker, R.; Sandur, S.K.; Jawali, N. Antioxidant supplementation enhances bacterial peritonitis in mice by inhibiting phagocytosis. J. Med. Microbiol. 2014, 63, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Orecchioni, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Macrophage Polarization: Different Gene Signatures in M1(LPS+) vs. Classically and M2(LPS-) vs. Alternatively Activated Macrophages. Front. Immunol. 2019, 10, 1084. [Google Scholar] [CrossRef] [PubMed]
- Ciesielska, A.; Matyjek, M.; Kwiatkowska, K. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell. Mol. Life Sci. 2021, 78, 1233–1261. [Google Scholar] [CrossRef] [PubMed]
- Reddy, C.S.; Kim, S.C.; Hur, M.; Kim, Y.B.; Park, C.G.; Lee, W.M.; Jang, J.K.; Koo, S.C. Natural Korean Medicine Dang-Gui: Biosynthesis, Effective Extraction and Formulations of Major Active Pyranocoumarins, Their Molecular Action Mechanism in Cancer, and Other Biological Activities. Molecules 2017, 22, 2170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canton, J. Phagosome maturation in polarized macrophages. J. Leukoc. Biol. 2014, 96, 729–738. [Google Scholar] [CrossRef]
- Jung, M.H.; Lee, S.H.; Ahn, E.M.; Lee, Y.M. Decursin and decursinol angelate inhibit VEGF-induced angiogenesis via suppression of the VEGFR-2-signaling pathway. Carcinogenesis 2009, 30, 655–661. [Google Scholar] [CrossRef] [Green Version]
- Vergadi, E.; Ieronymaki, E.; Lyroni, K.; Vaporidi, K.; Tsatsanis, C. Akt Signaling Pathway in Macrophage Activation and M1/M2 Polarization. J. Immunol. 2017, 198, 1006–1014. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Wang, Y.; Qin, Y.; He, W.; Benlahrech, A.; Zhang, Q.; Lu, Z.; Ji, G.; Zheng, Y. Micheliolide provides protection of mice against Staphylococcus aureus and MRSA infection by down-regulating inflammatory response. Sci. Rep. 2017, 7, 41964. [Google Scholar] [CrossRef] [Green Version]
- Pidwill, G.R.; Gibson, J.F.; Cole, J.; Renshaw, S.A.; Foster, S.J. The Role of Macrophages in Staphylococcus aureus Infection. Front. Immunol. 2020, 11, 620339. [Google Scholar] [CrossRef]
- Kim, K.M.; Kim, M.J.; Kang, J.S. Absorption, distribution, metabolism, and excretion of decursin and decursinol angelate from Angelica gigas Nakai. J. Microbiol. Biotechnol. 2009, 19, 1569–1572. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Lee, Y.S.; Jung, S.H.; Shin, K.H.; Kim, B.K.; Kang, S.S. Anti-tumor activities of decursinol angelate and decursin from Angelica gigas. Arch. Pharm. Res. 2003, 26, 727–730. [Google Scholar] [CrossRef]
- Kim, B.S.; Seo, H.; Kim, H.J.; Bae, S.M.; Son, H.N.; Lee, Y.J.; Ryu, S.; Park, R.W.; Nam, J.O. Decursin from Angelica gigas Nakai Inhibits B16F10 Melanoma Growth Through Induction of Apoptosis. J. Med. Food 2015, 18, 1121–1127. [Google Scholar] [CrossRef] [Green Version]
- Rose, W.; Fantl, M.; Geriak, M.; Nizet, V.; Sakoulas, G. Current Paradigms of Combination therapy in Methicillin-Resistant Staphylococcus aureus (MRSA) Bacteremia: Does it Work, Which Combination and For Which Patients? Clin. Infect. Dis. 2021, ciab452. [Google Scholar] [CrossRef] [PubMed]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Maharjan, S.; Park, B.K.; Lee, S.I.; Lim, Y.; Lee, K.; Kwon, H.J. Gomisin G Inhibits the Growth of Triple-Negative Breast Cancer Cells by Suppressing AKT Phosphorylation and Decreasing Cyclin D1. Biomol. Ther. 2018, 26, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, K.; Kim, M.S.; Ichikawa, R.; Ryu, K.H.; Dohmae, N.; Nakayama, H.; Lee, B.L. Environment-mediated accumulation of diacyl lipoproteins over their triacyl counterparts in Staphylococcus aureus. J. Bacteriol. 2012, 194, 3299–3306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.H.; Park, J.; Kim, D.; Gautam, A.; Akauliya, M.; Kim, J.; Lee, H.; Park, S.; Lee, Y.; Kwon, H.J. Anti-Bacterial Effect of CpG-DNA Involves Enhancement of the Complement Systems. Int. J. Mol. Sci. 2019, 20, 3397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sim, H.; Jeong, D.; Kim, H.I.; Pak, S.; Thapa, B.; Kwon, H.J.; Lee, K. CD11b Deficiency Exacerbates Methicillin-Resistant Staphylococcus aureus-Induced Sepsis by Upregulating Inflammatory Responses of Macrophages. Immune Netw. 2021, 21, e13. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Gu, Y.; Shin, A.; Zhang, S.; Ginhoux, F. Analysis of Myeloid Cells in Mouse Tissues with Flow Cytometry. STAR Protoc. 2020, 1, 100029. [Google Scholar] [CrossRef]
- Drevets, D.A.; Canono, B.P.; Campbell, P.A. Measurement of bacterial ingestion and killing by macrophages. Curr. Protoc. Immunol. 2015, 109, 14.6.1–14.6.17. [Google Scholar] [CrossRef]
- Herb, M.; Gluschko, A.; Wiegmann, K.; Farid, A.; Wolf, A.; Utermohlen, O.; Krut, O.; Kronke, M.; Schramm, M. Mitochondrial reactive oxygen species enable proinflammatory signaling through disulfide linkage of NEMO. Sci. Signal. 2019, 12, eaar5926. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pak, S.; Thapa, B.; Lee, K. Decursinol Angelate Mitigates Sepsis Induced by Methicillin-Resistant Staphylococcus aureus Infection by Modulating the Inflammatory Responses of Macrophages. Int. J. Mol. Sci. 2021, 22, 10950. https://doi.org/10.3390/ijms222010950
Pak S, Thapa B, Lee K. Decursinol Angelate Mitigates Sepsis Induced by Methicillin-Resistant Staphylococcus aureus Infection by Modulating the Inflammatory Responses of Macrophages. International Journal of Molecular Sciences. 2021; 22(20):10950. https://doi.org/10.3390/ijms222010950
Chicago/Turabian StylePak, Seongwon, Bikash Thapa, and Keunwook Lee. 2021. "Decursinol Angelate Mitigates Sepsis Induced by Methicillin-Resistant Staphylococcus aureus Infection by Modulating the Inflammatory Responses of Macrophages" International Journal of Molecular Sciences 22, no. 20: 10950. https://doi.org/10.3390/ijms222010950