
Curcumin Suppresses the Lipid Accumulation and Oxidative Stress Induced by Benzo[a]pyrene Toxicity in HepG2 Cells

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cell Culture
2.3. Cell Viability Assay
2.4. Assessment of Intracellular ROS Level
2.5. ORO Staining
2.6. Measurement of Triglyceride and Free-Glycerol Levels
2.7. Immunofluorescence Staining
2.8. siRNA Transfection
2.9. Western Blot Analysis
2.10. Statistical Analysis
3. Results
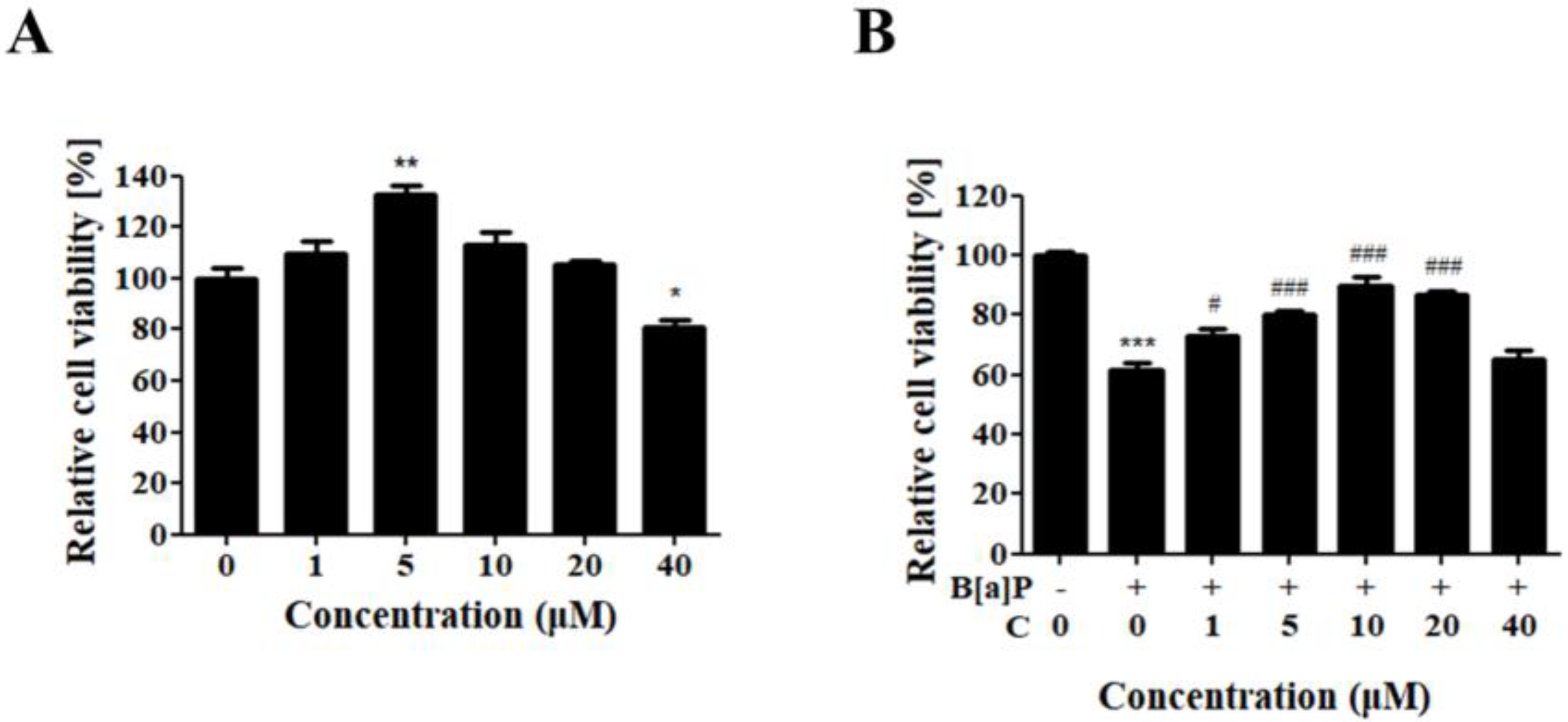
3.1. Protective Effect of Curcumin against B[a]P-Induced Cytotoxicity
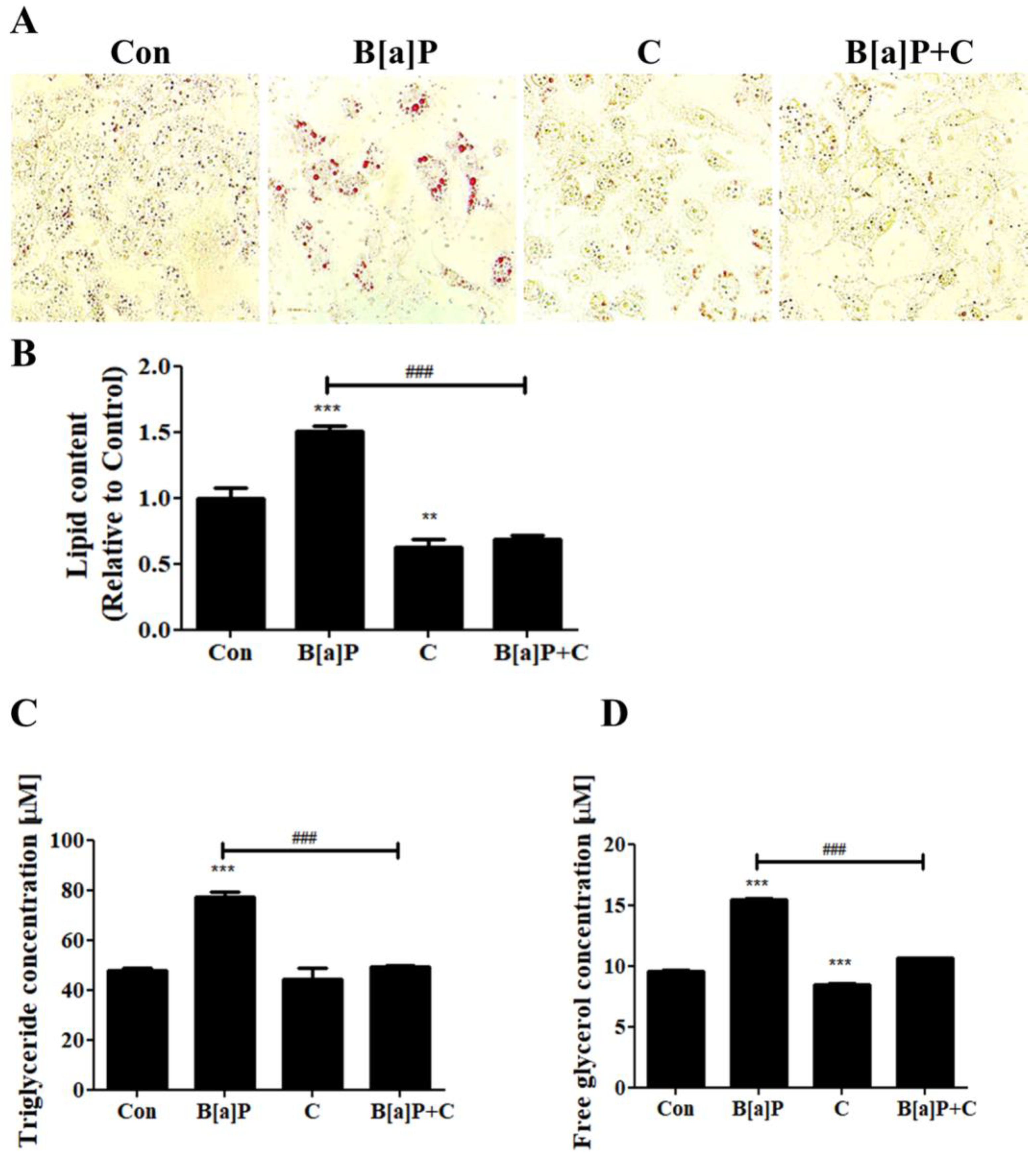
3.2. Suppressive Effect of Curcumin on the Lipid Accumulation Induced by B[a]P
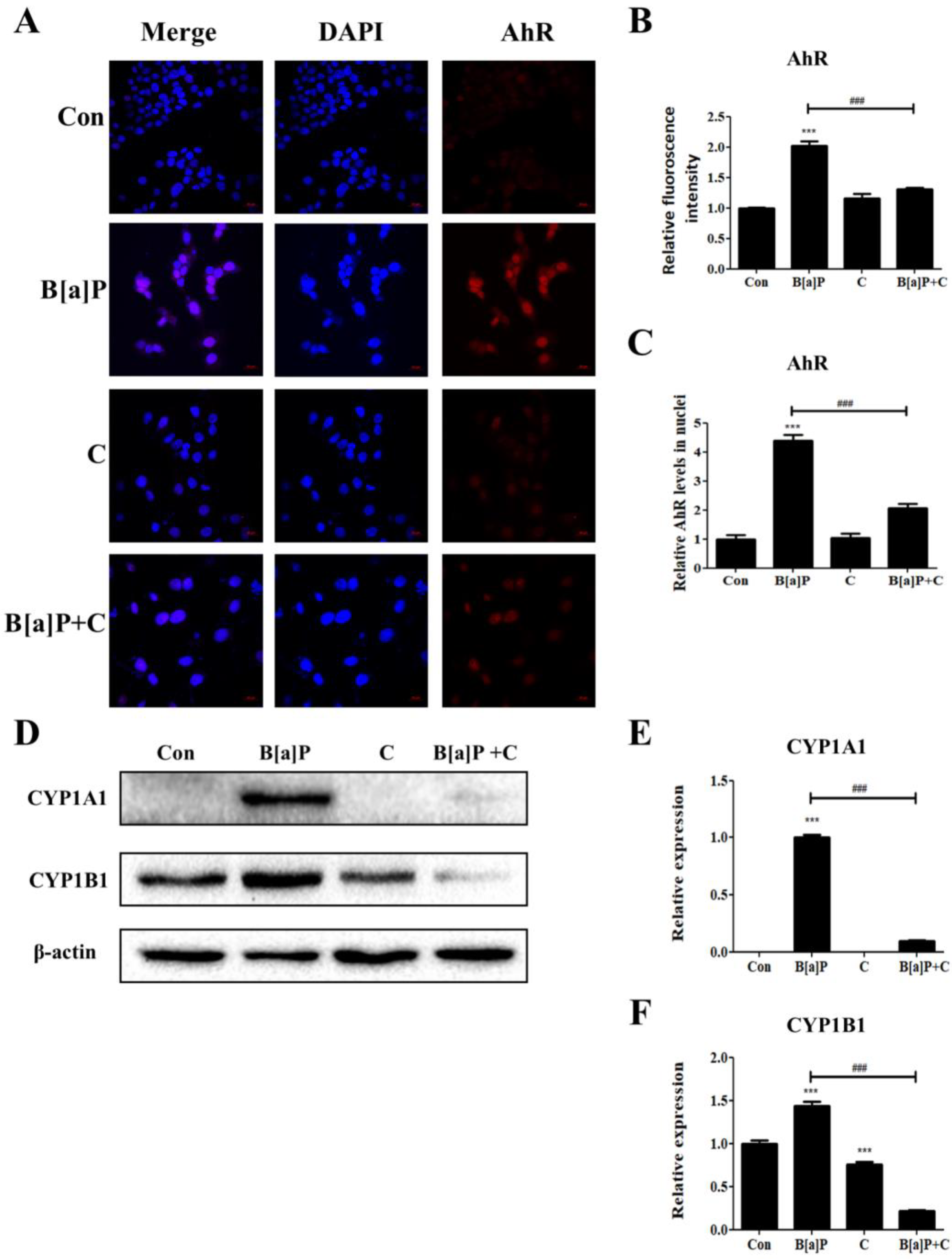
3.3. Inhibitory Effect of Curcumin on B[a]P-Induced AhR Pathways
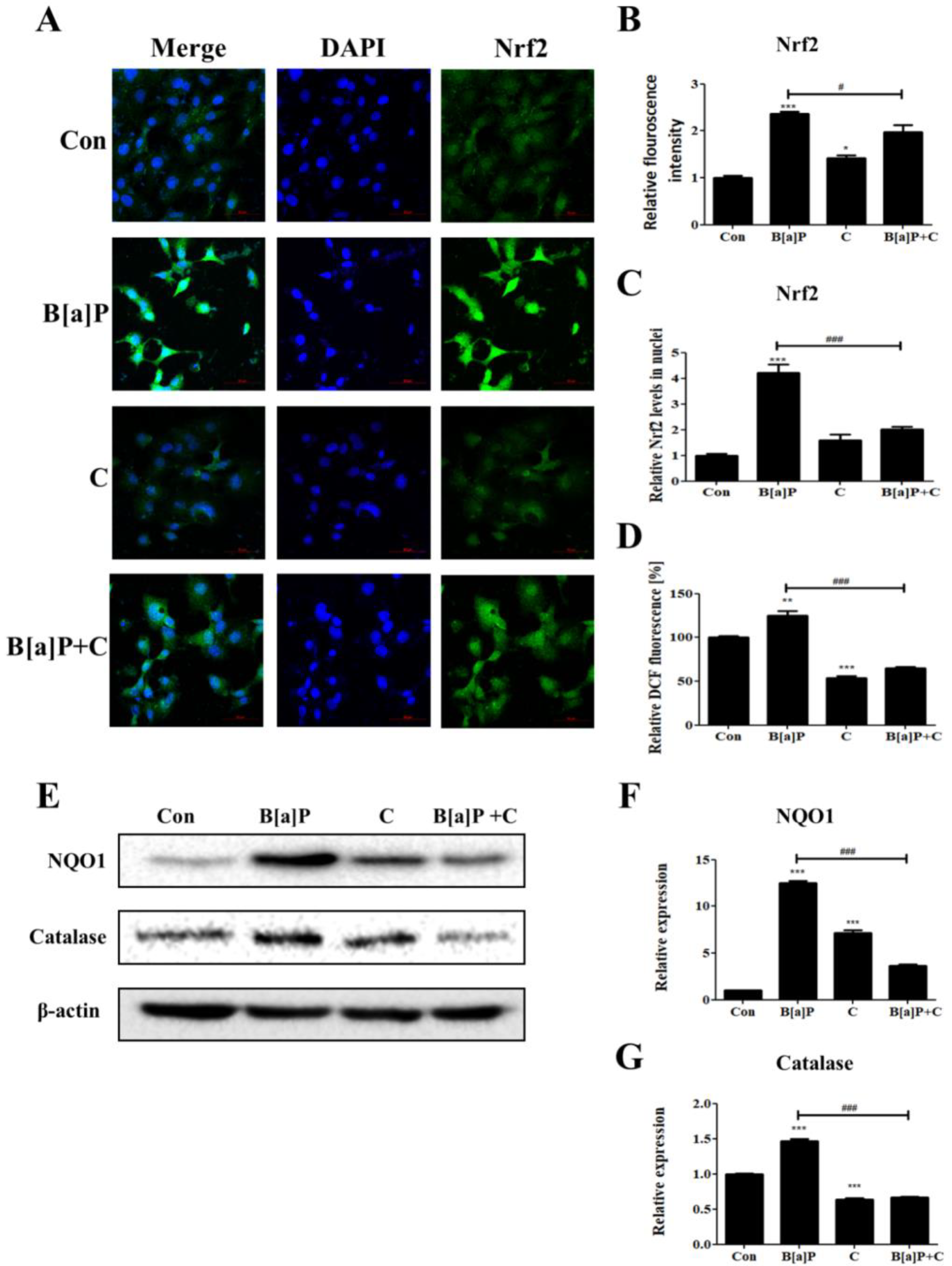
3.4. Inhibitory Effect of Curcumin on B[a]P-Induced Nrf2 Pathways
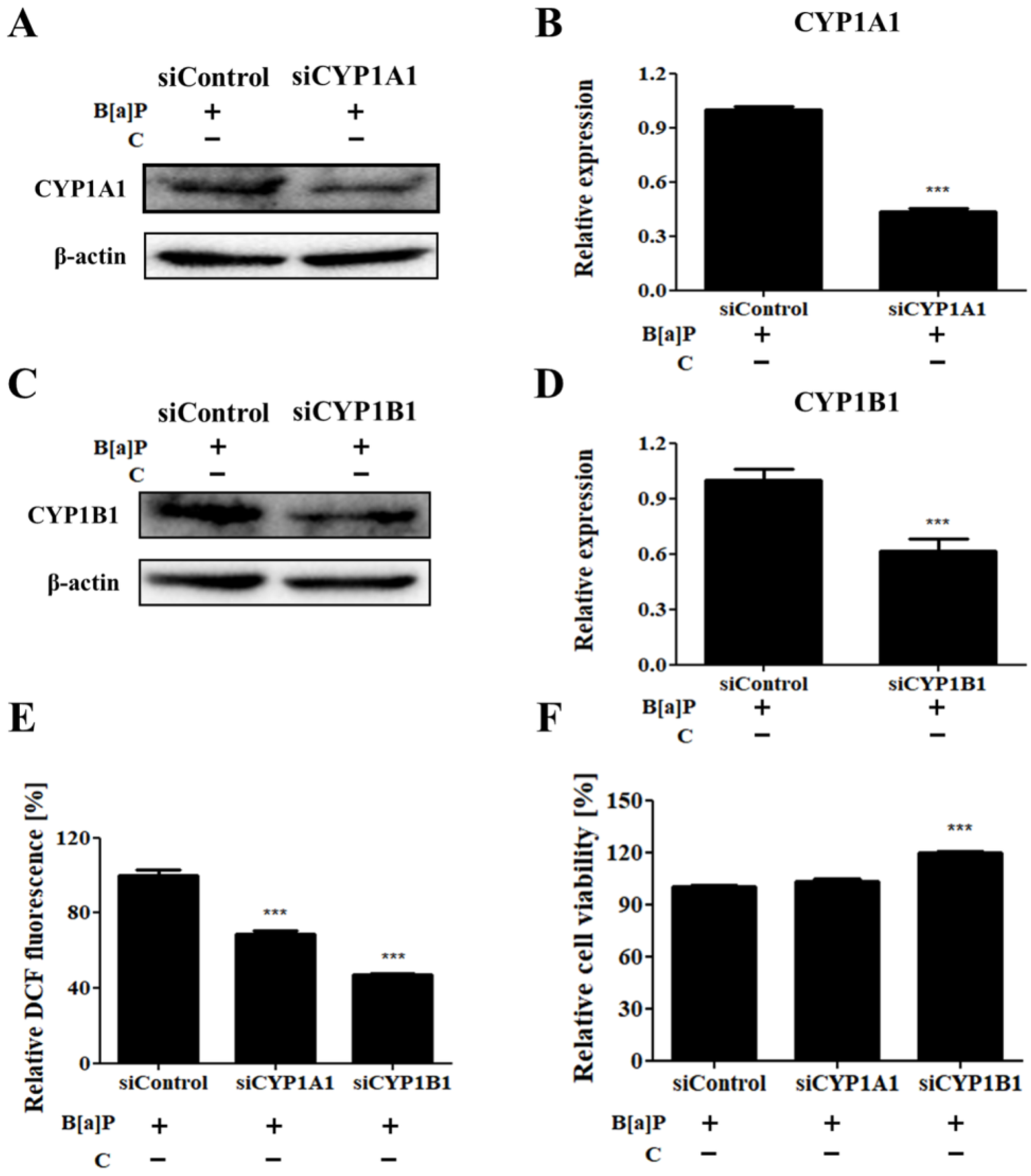
3.5. Effects of Cyp1a1 and Cyp1b1 Knockdown on B[a]P-Induced Cytotoxicity and Induction of ROS
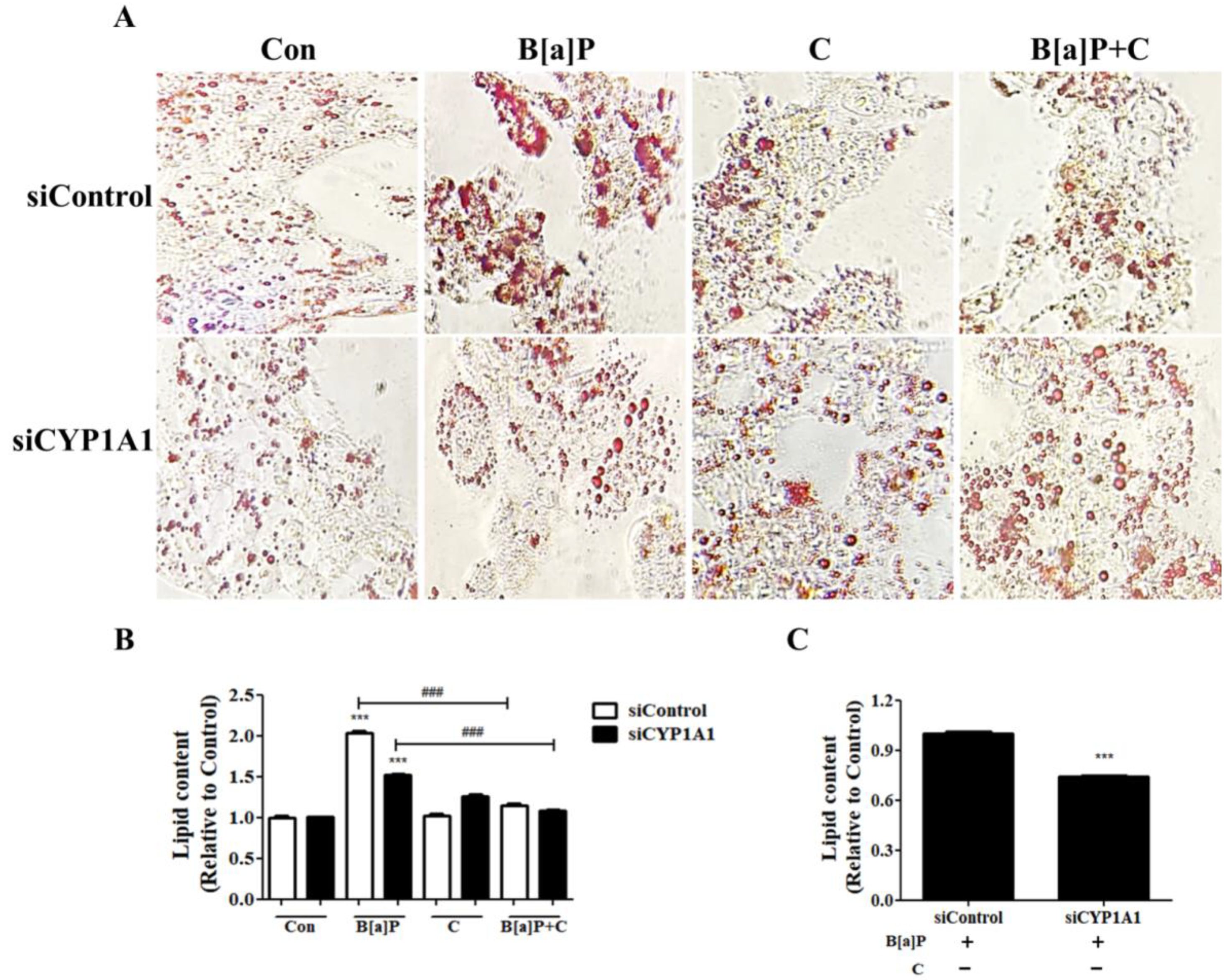
3.6. Effect of Cyp1a1 Knockdown on B[a]P-Induced Lipid Accumulation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ejaz, A.; Wu, D.; Kwan, P.; Meydani, M. Curcumin inhibits adipogenesis in 3T3-L1 adipocytes and angiogenesis and obesity in C57/BL mice. J. Nutr. 2009, 139, 919–925. [Google Scholar] [CrossRef]
- Aggarwal, B.B. Targeting Inflammation-Induced Obesity and Metabolic Diseases by Curcumin and Other Nutraceuticals. Annu. Rev. Nutr. 2010, 30, 173–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alappat, L.; Awad, A.B. Curcumin and obesity: Evidence and mechanisms. Nutr. Rev. 2010, 68, 729–738. [Google Scholar] [CrossRef]
- Kang, O.H.; Kim, S.B.; Seo, Y.S.; Joung, D.K.; Mun, S.H.; Choi, J.G.; Lee, Y.M.; Kang, D.G.; Lee, H.S.; Kwon, D.Y. Curcumin decreases oleic acid-induced lipid accumulation via AMPK phosphorylation in hepatocarcinoma cells. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 2578–2586. [Google Scholar]
- White, C.M.; Lee, J.Y. The impact of turmeric or its curcumin extract on nonalcoholic fatty liver disease: A systematic review of clinical trials. Pharm. Pract. 2019, 17, 1350. [Google Scholar] [CrossRef] [Green Version]
- Cicero, A.F.G.; Sahebkar, A.; Fogacci, F.; Bove, M.; Giovannini, M.; Borghi, C. Effects of phytosomal curcumin on anthropometric parameters, insulin resistance, cortisolemia and non-alcoholic fatty liver disease indices: A double-blind, placebo-controlled clinical trial. Eur. J. Nutr. 2020, 59, 477–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jee, S.C.; Kim, M.; Kim, K.S.; Kim, H.S.; Sung, J.S. Protective Effects of Myricetin on Benzo[a]pyrene-Induced 8-Hydroxy-2′-Deoxyguanosine and BPDE-DNA Adduct. Antioxidants 2020, 9, 446. [Google Scholar] [CrossRef]
- Jee, S.C.; Kim, M.; Sung, J.S. Modulatory Effects of Silymarin on Benzo[a]pyrene-Induced Hepatotoxicity. Int. J. Mol. Sci. 2020, 21, 2369. [Google Scholar] [CrossRef] [Green Version]
- Larsson, B.K.; Sahlberg, G.P.; Eriksson, A.T.; Busk, L.A. Polycyclic aromatic hydrocarbons in grilled food. J. Agric. Food Chem. 1983, 31, 867–873. [Google Scholar] [CrossRef]
- Nisbet, I.C.T.; LaGoy, P.K. Toxic equivalency factors (TEFs) for polycyclic aromatic hydrocarbons (PAHs). Regul. Toxicol. Pharmacol. 1992, 16, 290–300. [Google Scholar] [CrossRef]
- Baan, R.; Grosse, Y.; Straif, K.; Secretan, B.; El Ghissassi, F.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Freeman, C.; Galichet, L.; et al. A review of human carcinogens--Part F: Chemical agents and related occupations. Lancet Oncol. 2009, 10, 1143–1144. [Google Scholar] [CrossRef]
- Perera, F.; Tang, D.; Whyatt, R.; Lederman, S.A.; Jedrychowski, W. DNA damage from polycyclic aromatic hydrocarbons measured by benzo[a]pyrene-DNA adducts in mothers and newborns from Northern Manhattan, the World Trade Center Area, Poland, and China. Cancer Epidemiol. Biomark. Prev. 2005, 14, 709–714. [Google Scholar] [CrossRef] [Green Version]
- Hanelt, S.; Helbig, R.; Hartmann, A.; Lang, M.; Seidel, A.; Speit, G. A comparative investigation of DNA adducts, DNA strand breaks and gene mutations induced by benzo[a]pyrene and (+/−)-anti-benzo[a]pyrene-7,8-diol 9,10-oxide in cultured human cells. Mutat. Res. 1997, 390, 179–188. [Google Scholar] [CrossRef]
- Boysen, G.; Hecht, S.S. Analysis of DNA and protein adducts of benzo[a]pyrene in human tissues using structure-specific methods. Mutat. Res. 2003, 543, 17–30. [Google Scholar] [CrossRef]
- Walle, T.; Walle, U.K.; Sedmera, D.; Klausner, M. Benzo[A]pyrene-induced oral carcinogenesis and chemoprevention: Studies in bioengineered human tissue. Drug Metab. Dispos. 2006, 34, 346–350. [Google Scholar] [CrossRef] [PubMed]
- Ueng, T.H.; Chang, Y.L.; Tsai, Y.Y.; Su, J.L.; Chan, P.K.; Shih, J.Y.; Lee, Y.C.; Ma, Y.C.; Kuo, M.L. Potential roles of fibroblast growth factor-9 in the benzo(a)pyrene-induced invasion in vitro and the metastasis of human lung adenocarcinoma. Arch. Toxicol. 2010, 84, 651–660. [Google Scholar] [CrossRef]
- Li, D.; Firozi, P.F.; Wang, L.E.; Bosken, C.H.; Spitz, M.R.; Hong, W.K.; Wei, Q. Sensitivity to DNA damage induced by benzo(a)pyrene diol epoxide and risk of lung cancer: A case-control analysis. Cancer Res. 2001, 61, 1445–1450. [Google Scholar]
- Widziewicz, K.; Rogula-Kozłowska, W.; Majewski, G. Lung Cancer Risk Associated with Exposure to Benzo(A)Pyrene in Polish Agglomerations, Cities, and Other Areas. Int. J. Environ. Res. 2017, 11, 685–693. [Google Scholar] [CrossRef] [Green Version]
- Amadou, A.; Praud, D.; Coudon, T.; Deygas, F.; Grassot, L.; Faure, E.; Couvidat, F.; Caudeville, J.; Bessagnet, B.; Salizzoni, P.; et al. Risk of breast cancer associated with long-term exposure to benzo[a]pyrene (BaP) air pollution: Evidence from the French E3N cohort study. Environ. Int. 2021, 149, 106399. [Google Scholar] [CrossRef]
- Defois, C.; Ratel, J.; Denis, S.; Batut, B.; Beugnot, R.; Peyretaillade, E.; Engel, E.; Peyret, P. Environmental Pollutant Benzo[a]Pyrene Impacts the Volatile Metabolome and Transcriptome of the Human Gut Microbiota. Front. Microbiol. 2017, 8, 1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Xu, Y.; Ji, W.; Song, L.; Dai, C.; Zhan, L. Effects of exposure to benzo[a]pyrene on metastasis of breast cancer are mediated through ROS-ERK-MMP9 axis signaling. Toxicol. Lett. 2015, 234, 201–210. [Google Scholar] [CrossRef]
- Ba, Q.; Li, J.; Huang, C.; Qiu, H.; Li, J.; Chu, R.; Zhang, W.; Xie, D.; Wu, Y.; Wang, H. Effects of benzo[a]pyrene exposure on human hepatocellular carcinoma cell angiogenesis, metastasis, and NF-κB signaling. Environ. Health Perspect. 2015, 123, 246–254. [Google Scholar] [CrossRef] [Green Version]
- Steck, S.E.; Gaudet, M.M.; Eng, S.M.; Britton, J.A.; Teitelbaum, S.L.; Neugut, A.I.; Santella, R.M.; Gammon, M.D. Cooked meat and risk of breast cancer--lifetime versus recent dietary intake. Epidemiology 2007, 18, 373–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.G.; Burstyn, I.; Lai, A.S.; Grundy, A.; Friesen, M.C.; Aronson, K.J.; Spinelli, J.J. Women’s occupational exposure to polycyclic aromatic hydrocarbons and risk of breast cancer. Occup. Environ. Med. 2019, 76, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Zhao, B.; Zhang, J.; Martin, F.L.; Huang, Q.; Liu, L.; Shen, H. Association of environmental benzo[a]pyrene exposure and DNA methylation alterations in hepatocellular carcinoma: A Chinese case-control study. Sci. Total Environ. 2016, 541, 1243–1252. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.; Zhao, B.; Guo, F.; Bin, Z.; Yang, Y.; Liu, S.; Han, Y.; Niu, J.; Ke, X.; Wang, N.; et al. Interaction of benzo[a]pyrene with other risk factors in hepatocellular carcinoma: A case-control study in Xiamen, China. Ann. Epidemiol. 2014, 24, 98–103. [Google Scholar] [CrossRef]
- Ortiz, L.; Nakamura, B.; Li, X.; Blumberg, B.; Luderer, U. In utero exposure to benzo[a]pyrene increases adiposity and causes hepatic steatosis in female mice, and glutathione deficiency is protective. Toxicol. Lett. 2013, 223, 260–267. [Google Scholar] [CrossRef] [Green Version]
- Ramesh, A.; Hood, D.B.; Inyang, F.; Greenwood, M.; Nyanda, A.M.; Archibong, A.E.; Knuckles, M.E. Comparative metabolism, bioavailability, and toxicokinetics of benzo[a]pyrene in rats after acute oral, inhalation, and intravenous administration. Polycycl. Aromat. Comp. 2002, 22, 969–980. [Google Scholar] [CrossRef]
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global Epidemiology of Nonalcoholic Fatty Liver Disease-Meta-Analytic Assessment of Prevalence, Incidence, and Outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, C.R.; Nguyen, M.H.; Lim, J.K. Hepatocellular carcinoma in patients with non-alcoholic fatty liver disease. World J. Gastroentero 2016, 22, 8294–8303. [Google Scholar] [CrossRef]
- Siegel, A.B.; Zhu, A.X. Metabolic Syndrome and Hepatocellular Carcinoma Two Growing Epidemics With a Potential Link. Cancer-Am. Cancer Soc. 2009, 115, 5651–5661. [Google Scholar] [CrossRef] [Green Version]
- Harper, P.A.; Prokipcak, R.D.; Bush, L.E.; Golas, C.L.; Okey, A.B. Detection and characterization of the Ah receptor for 2,3,7,8-tetrachlorodibenzo-p-dioxin in the human colon adenocarcinoma cell line LS180. Arch. Biochem. Biophys. 1991, 290, 27–36. [Google Scholar] [CrossRef]
- Fukunaga, B.N.; Probst, M.R.; Reisz-Porszasz, S.; Hankinson, O. Identification of functional domains of the aryl hydrocarbon receptor. J. Biol. Chem. 1995, 270, 29270–29278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, G.I.; Melvin, W.T.; Greenlee, W.F.; Burke, M.D. Regulation, function, and tissue-specific expression of cytochrome P450 CYP1B1. Annu. Rev. Pharmacol. Toxicol. 2001, 41, 297–316. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.S.S.; Ande, A.; Sinha, N.; Kumar, A.; Kumar, S. Effects of Cigarette Smoke Condensate on Oxidative Stress, Apoptotic Cell Death, and HIV Replication in Human Monocytic Cells. PLoS ONE 2016, 11, e0155791. [Google Scholar] [CrossRef] [Green Version]
- Veith, A.; Moorthy, B. Role of cytochrome p450s in the generation and metabolism of reactive oxygen species. Curr. Opin. Toxicol. 2018, 7, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Bao, J.; Cao, Y.R.; Gao, H.F.; Jin, Y. Cytochrome P450 1A1 (CYP1A1) Catalyzes Lipid Peroxidation of Oleic Acid-Induced HepG2 Cells. Biochemistry 2018, 83, 595–602. [Google Scholar] [CrossRef]
- Filip, R.; Shaw, T.A.; Nishida, A.; Pezacki, J.P. Fungal natural alkaloid schizocommunin activates the aryl hydrocarbon receptor pathway. Medchemcomm 2019, 10, 985–990. [Google Scholar] [CrossRef]
- Ohashi, H.; Nishioka, K.; Nakajima, S.; Kim, S.; Suzuki, R.; Aizaki, H.; Fukasawa, M.; Kamisuki, S.; Sugawara, F.; Ohtani, N.; et al. The aryl hydrocarbon receptor-cytochrome P450 1A1 pathway controls lipid accumulation and enhances the permissiveness for hepatitis C virus assembly. J. Biol. Chem. 2018, 293, 19559–19571. [Google Scholar] [CrossRef] [Green Version]
- Park, C.G.; Sung, B.; Ryu, C.S.; Kim, Y.J. Mono-(2-ethylhexyl) phthalate induces oxidative stress and lipid accumulation in zebrafish liver cells. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2020, 230, 108704. [Google Scholar] [CrossRef]
- Shimizu, Y.; Nakatsuru, Y.; Ichinose, M.; Takahashi, Y.; Kume, H.; Mimura, J.; Fujii-Kuriyama, Y.; Ishikawa, T. Benzo[a]pyrene carcinogenicity is lost in mice lacking the aryl hydrocarbon receptor. Proc. Natl. Acad. Sci. USA 2000, 97, 779–782. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q. Role of nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, S.; Wakabayashi, N.; Misra, V.; Biswal, S.; Lee, G.H.; Agoston, E.S.; Yamamoto, M.; Kensler, T.W. NRF2 modulates aryl hydrocarbon receptor signaling: Influence on adipogenesis. Mol. Cell Biol. 2007, 27, 7188–7197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantey, J.A.; Rekhadevi, P.V.; Diggs, D.L.; Ramesh, A. Metabolism of benzo(a)pyrene by subcellular fractions of gastrointestinal (GI) tract and liver in Apc(Min) mouse model of colon cancer. Tumour. Biol. 2014, 35, 4929–4935. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Rayess, Y.E.; Rizk, A.A.; Sadaka, C.; Zgheib, R.; Zam, W.; Sestito, S.; Rapposelli, S.; Neffe-Skocinska, K.; Zielinska, D.; et al. Turmeric and Its Major Compound Curcumin on Health: Bioactive Effects and Safety Profiles for Food, Pharmaceutical, Biotechnological and Medicinal Applications. Front. Pharmacol. 2020, 11, 01021. [Google Scholar] [CrossRef] [PubMed]
- Menniti-Ippolito, F.; Ippoliti, I.; Pastorelli, A.A.; Altieri, I.; Scalise, F.; De Santis, B.; Debegnach, F.; Brera, C.; Pacifici, R.; Pichini, S.; et al. Turmeric (Curcuma longa L.) food supplements and hepatotoxicity: An integrated evaluation approach. Ann. Ist Super Sanita 2020, 56, 462–469. [Google Scholar] [CrossRef]
- Kocaadam, B.; Sanlier, N. Curcumin, an active component of turmeric (Curcuma longa), and its effects on health. Crit. Rev. Food Sci. Nutr. 2017, 57, 2889–2895. [Google Scholar] [CrossRef]
- Kim, K.S.; Kim, N.Y.; Son, J.Y.; Park, J.H.; Lee, S.H.; Kim, H.R.; Kim, B.; Kim, Y.G.; Jeong, H.G.; Lee, B.M.; et al. Curcumin Ameliorates Benzo[a]pyrene-Induced DNA Damages in Stomach Tissues of Sprague-Dawley Rats. Int. J. Mol. Sci. 2019, 20, 5533. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wu, Y.M.; Zhang, P.Y. Protective effects of curcumin and quercetin during benzo(a)pyrene induced lung carcinogenesis in mice. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 1736–1743. [Google Scholar]
- Chen, Z.; Tian, R.; She, Z.; Cai, J.; Li, H. Role of oxidative stress in the pathogenesis of nonalcoholic fatty liver disease. Free Radic Biol. Med. 2020, 152, 116–141. [Google Scholar] [CrossRef] [PubMed]
- Sekiya, M.; Hiraishi, A.; Touyama, M.; Sakamoto, K. Oxidative stress induced lipid accumulation via SREBP1c activation in HepG2 cells. Biochem. Biophys. Res. Commun. 2008, 375, 602–607. [Google Scholar] [CrossRef] [Green Version]
- Nebert, D.W.; Roe, A.L.; Dieter, M.Z.; Solis, W.A.; Yang, Y.; Dalton, T.P. Role of the aromatic hydrocarbon receptor and [Ah] gene battery in the oxidative stress response, cell cycle control, and apoptosis. Biochem. Pharmacol. 2000, 59, 65–85. [Google Scholar] [CrossRef]
- Garg, R.; Gupta, S.; Maru, G.B. Dietary curcumin modulates transcriptional regulators of phase I and phase II enzymes in benzo[a]pyrene-treated mice: Mechanism of its anti-initiating action. Carcinogenesis 2008, 29, 1022–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakai, R.; Fukuda, S.; Kawase, M.; Yamashita, Y.; Ashida, H. Curcumin and its derivatives inhibit 2,3,7,8,-tetrachloro-dibenzo-p-dioxin-induced expression of drug metabolizing enzymes through aryl hydrocarbon receptor-mediated pathway. Biosci. Biotechnol. Biochem. 2018, 82, 616–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional Regulation by Nrf2. Antioxid. Redox Signal. 2018, 29, 1727–1745. [Google Scholar] [CrossRef] [Green Version]
- Neilson, L.E.; Quinn, J.F.; Gray, N.E. Peripheral Blood NRF2 Expression as a Biomarker in Human Health and Disease. Antioxidants 2020, 10, 28. [Google Scholar] [CrossRef]
- Burczynski, M.E.; Penning, T.M. Genotoxic polycyclic aromatic hydrocarbon ortho-quinones generated by aldo-keto reductases induce CYP1A1 via nuclear translocation of the aryl hydrocarbon receptor. Cancer Res. 2000, 60, 908–915. [Google Scholar]
- Vorrink, S.U.; Hudachek, D.R.; Domann, F.E. Epigenetic determinants of CYP1A1 induction by the aryl hydrocarbon receptor agonist 3,3′,4,4′,5-pentachlorobiphenyl (PCB 126). Int. J. Mol. Sci. 2014, 15, 13916–13931. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-C.; Jee, S.-C.; Kim, M.; Kim, S.; Shin, M.K.; Kim, Y.; Sung, J.-S. Curcumin Suppresses the Lipid Accumulation and Oxidative Stress Induced by Benzo[a]pyrene Toxicity in HepG2 Cells. Antioxidants 2021, 10, 1314. https://doi.org/10.3390/antiox10081314
Lee S-C, Jee S-C, Kim M, Kim S, Shin MK, Kim Y, Sung J-S. Curcumin Suppresses the Lipid Accumulation and Oxidative Stress Induced by Benzo[a]pyrene Toxicity in HepG2 Cells. Antioxidants. 2021; 10(8):1314. https://doi.org/10.3390/antiox10081314
Chicago/Turabian StyleLee, Seung-Cheol, Seung-Cheol Jee, Min Kim, Soee Kim, Min Kyoung Shin, Yunkyung Kim, and Jung-Suk Sung. 2021. "Curcumin Suppresses the Lipid Accumulation and Oxidative Stress Induced by Benzo[a]pyrene Toxicity in HepG2 Cells" Antioxidants 10, no. 8: 1314. https://doi.org/10.3390/antiox10081314